- ไมโตคอนเดรียเป็นการรวมตัวกันเล็กๆ ในเซลล์ที่แต่เดิมคิดว่าสืบทอดมาจากแบคทีเรีย ในเซลล์ส่วนใหญ่มีมากถึงหลายพันเซลล์ ซึ่งคิดเป็นตั้งแต่ 15 ถึง 50 เปอร์เซ็นต์ของปริมาตรเซลล์ เป็นแหล่งพลังงานมากกว่า 90 เปอร์เซ็นต์ของร่างกายของคุณ
- ไมโตคอนเดรียของคุณส่งผลกระทบอย่างมากต่อสุขภาพ โดยเฉพาะมะเร็ง ดังนั้นการปรับปรุงการเผาผลาญของไมโตคอนเดรียอาจเป็นหัวใจสำคัญของการรักษามะเร็งที่มีประสิทธิผล
ขนาดข้อความ:
จาก ดร.เมอร์โคลา
Mitochondria: คุณอาจไม่รู้ว่ามันคืออะไร แต่มันคืออะไร สำคัญยิ่งเพื่อสุขภาพของคุณ Rhonda Patrick ปริญญาเอกเป็นนักวิทยาศาสตร์ชีวการแพทย์ที่ได้ศึกษาปฏิสัมพันธ์ระหว่างเมแทบอลิซึมของไมโตคอนเดรีย เมแทบอลิซึมที่ผิดปกติ และมะเร็ง
งานส่วนหนึ่งของเธอเกี่ยวข้องกับการระบุตัวบ่งชี้ทางชีวภาพของโรคในระยะเริ่มแรก ตัวอย่างเช่น ความเสียหายของ DNA ถือเป็นตัวบ่งชี้ทางชีวภาพระยะแรกของมะเร็ง จากนั้นเธอก็พยายามพิจารณาว่าสารอาหารรองชนิดใดที่ช่วยซ่อมแซมความเสียหายของ DNA นี้
เธอยังค้นคว้าเกี่ยวกับการทำงานของไมโตคอนเดรียและเมแทบอลิซึม ซึ่งเป็นสิ่งที่ฉันสนใจเมื่อเร็วๆ นี้ หลังจากฟังการสัมภาษณ์นี้แล้ว หากคุณต้องการเรียนรู้เพิ่มเติมเกี่ยวกับเรื่องนี้ ฉันขอแนะนำให้เริ่มต้นด้วยหนังสือ Life - The Epic Story of Our Mitochondria ของ Dr. Lee Know
ไมโตคอนเดรียมีผลกระทบอย่างมากต่อสุขภาพ โดยเฉพาะมะเร็ง และฉันเริ่มเชื่อว่าการเพิ่มประสิทธิภาพการเผาผลาญของไมโตคอนเดรียอาจเป็นหัวใจสำคัญของการรักษาโรคมะเร็งที่มีประสิทธิผล
ความสำคัญของการเพิ่มประสิทธิภาพการเผาผลาญของไมโตคอนเดรีย
ไมโตคอนเดรียเป็นออร์แกเนลล์เล็กๆ ที่เราเชื่อกันว่าสืบทอดมาจากแบคทีเรีย แทบไม่มีเลยในเซลล์เม็ดเลือดแดงและเซลล์ผิวหนัง แต่ในเซลล์สืบพันธุ์มี 100,000 เซลล์ แต่ในเซลล์ส่วนใหญ่มีตั้งแต่ 1 ถึง 2,000 เซลล์เหล่านี้เป็นแหล่งพลังงานหลักสำหรับร่างกายของคุณ
เพื่อให้อวัยวะต่างๆ ทำงานได้อย่างถูกต้อง พวกมันต้องการพลังงาน และพลังงานนี้ผลิตโดยไมโตคอนเดรีย
เนื่องจากการทำงานของไมโตคอนเดรียเป็นรากฐานของทุกสิ่งที่เกิดขึ้นในร่างกาย การเพิ่มประสิทธิภาพการทำงานของไมโตคอนเดรีย และการป้องกันความผิดปกติของไมโตคอนเดรียโดยการได้รับสารอาหารและสารตั้งต้นที่จำเป็นทั้งหมดที่จำเป็นสำหรับไมโตคอนเดรีย จึงมีความสำคัญอย่างยิ่งต่อสุขภาพและการป้องกันโรค
ดังนั้นลักษณะสากลอย่างหนึ่งของเซลล์มะเร็งคือการด้อยค่าอย่างร้ายแรงของการทำงานของไมโตคอนเดรีย ซึ่งจำนวนไมโตคอนเดรียที่ทำงานได้ลดลงอย่างมาก
ดร. ออตโต วอร์เบิร์ก เป็นแพทย์ที่สำเร็จการศึกษาสาขาเคมีและเป็นเพื่อนสนิทของอัลเบิร์ต ไอน์สไตน์ ผู้เชี่ยวชาญส่วนใหญ่ยอมรับว่า Warburg เป็นนักชีวเคมีที่ยิ่งใหญ่ที่สุดแห่งศตวรรษที่ 20
ในปี 1931 เขาได้รับรางวัลโนเบลจากการค้นพบว่าเซลล์มะเร็งใช้กลูโคสเป็นแหล่งผลิตพลังงาน สิ่งนี้เรียกว่า “เอฟเฟกต์ Warburg” แต่น่าเสียดายที่ปรากฏการณ์นี้ยังคงถูกมองข้ามโดยเกือบทุกคน
ฉันเชื่อว่าการคุมอาหารแบบคีโตเจนิกซึ่งช่วยปรับปรุงสุขภาพของไมโตคอนเดรียได้อย่างมาก สามารถช่วยเหลือมะเร็งส่วนใหญ่ได้ โดยเฉพาะอย่างยิ่งเมื่อใช้ร่วมกับสารกำจัดกลูโคส เช่น 3-โบรโมไพรูเวท
ไมโตคอนเดรียผลิตพลังงานได้อย่างไร
ในการผลิตพลังงาน ไมโตคอนเดรียต้องการออกซิเจนจากอากาศที่คุณหายใจ และไขมันและกลูโคสจากอาหารที่คุณกิน
กระบวนการทั้งสองนี้ - การหายใจและการรับประทานอาหาร - เชื่อมต่อกันในกระบวนการที่เรียกว่าออกซิเดชั่นฟอสโฟรีเลชั่น ไมโตคอนเดรียใช้ในการผลิตพลังงานในรูปของเอทีพี
ไมโตคอนเดรียมีสายโซ่ขนส่งอิเล็กตรอนหลายชุด โดยพวกมันจะถ่ายโอนอิเล็กตรอนจากอาหารรูปแบบรีดิวซ์ที่คุณกินไปรวมกับออกซิเจนจากอากาศที่คุณหายใจจนกลายเป็นน้ำในที่สุด
กระบวนการนี้ขับเคลื่อนโปรตอนผ่านเยื่อหุ้มไมโตคอนเดรีย และชาร์จ ATP (adenosine triฟอสเฟต) จาก ADP (adenosine diฟอสเฟต) อีกครั้ง ATP ลำเลียงพลังงานไปทั่วร่างกาย
แต่กระบวนการนี้ทำให้เกิดผลพลอยได้ เช่น ออกซิเจนชนิดรีแอกทีฟ (ROS) ซึ่ง ความเสียหายเซลล์และ DNA ของไมโตคอนเดรีย จากนั้นจึงถ่ายโอนไปยัง DNA ของนิวเคลียส
การประนีประนอมจึงเกิดขึ้น โดยการผลิตพลังงานให้กับร่างกาย เริ่มแก่แล้วเนื่องจากลักษณะการทำลายล้างของ ROS ที่เกิดขึ้นในกระบวนการ อัตราที่ร่างกายมีอายุมากขึ้นนั้นขึ้นอยู่กับการทำงานของไมโตคอนเดรียเป็นหลักและปริมาณความเสียหายที่สามารถชดเชยได้ด้วยการปรับอาหารให้เหมาะสม
บทบาทของไมโตคอนเดรียต่อมะเร็ง
เมื่อเซลล์มะเร็งปรากฏขึ้น สายพันธุ์ออกซิเจนที่เกิดปฏิกิริยาซึ่งเป็นผลพลอยได้จากการผลิต ATP จะส่งสัญญาณที่กระตุ้นให้เกิดกระบวนการฆ่าตัวตายของเซลล์ หรือที่เรียกว่าอะพอพโทซิส
เนื่องจากเซลล์มะเร็งเกิดขึ้นทุกวัน นี่จึงเป็นสิ่งที่ดี โดยการฆ่าเซลล์ที่เสียหาย ร่างกายจะกำจัดเซลล์เหล่านั้นและแทนที่ด้วยเซลล์ที่มีสุขภาพดี
อย่างไรก็ตาม เซลล์มะเร็งสามารถต้านทานแผนการฆ่าตัวตายนี้ได้ พวกมันมีการป้องกันในตัว ดังที่ดร. วอร์เบิร์ก อธิบายและต่อมาโดยโธมัส ไซฟรีด ผู้ค้นคว้าวิจัยอย่างลึกซึ้งเกี่ยวกับมะเร็งว่าเป็นโรคทางเมตาบอลิซึม
ดังที่แพทริคอธิบาย:
“กลไกการออกฤทธิ์ประการหนึ่งของยาเคมีบำบัดคือการก่อตัวของสายพันธุ์ออกซิเจนที่เกิดปฏิกิริยา พวกมันสร้างความเสียหาย และนี่ก็เพียงพอแล้วที่จะผลักดันเซลล์มะเร็งไปสู่ความตาย
ฉันคิดว่าเหตุผลก็คือ เซลล์มะเร็งที่ไม่ได้ใช้ไมโตคอนเดรียของมัน กล่าวคือ ไม่ผลิตออกซิเจนชนิดที่มีปฏิกิริยาอีกต่อไป และจู่ๆ คุณก็บังคับให้มันใช้ไมโตคอนเดรีย และคุณมีออกซิเจนชนิดปฏิกิริยาเพิ่มขึ้นอย่างรวดเร็ว (ท้ายที่สุด นั่นคือสิ่งที่ไมโตคอนเดรียทำ) และ - บูม ความตาย เพราะเซลล์มะเร็งพร้อมแล้วสำหรับความตายครั้งนี้ เธอพร้อมที่จะตายแล้ว”
ทำไมไม่กินข้าวตอนเย็นถึงดี?
ฉันเป็นแฟนตัวยงของการอดอาหารไม่สม่ำเสมอมาระยะหนึ่งแล้วด้วยเหตุผลหลายประการ แน่นอนว่าการมีอายุยืนยาวและความกังวลเรื่องสุขภาพ แต่ยังเป็นเพราะดูเหมือนว่าจะให้ประโยชน์ในการป้องกันและรักษาโรคมะเร็งที่มีประสิทธิภาพ และกลไกนี้เกี่ยวข้องกับผลกระทบที่การอดอาหารมีต่อไมโตคอนเดรีย
ดังที่ได้กล่าวไปแล้ว ผลข้างเคียงที่สำคัญของการถ่ายโอนอิเล็กตรอนที่ไมโตคอนเดรียมีส่วนร่วมก็คือ บางส่วนรั่วไหลออกจากห่วงโซ่การขนส่งอิเล็กตรอนและทำปฏิกิริยากับออกซิเจนเพื่อสร้างอนุมูลอิสระซูเปอร์ออกไซด์
ซูเปอร์ออกไซด์ไอออน (ผลของการลดออกซิเจนด้วยอิเล็กตรอนหนึ่งตัว) เป็นสารตั้งต้นของออกซิเจนชนิดที่เกิดปฏิกิริยาส่วนใหญ่และเป็นสื่อกลางของปฏิกิริยาลูกโซ่ออกซิเดชั่น อนุมูลอิสระออกซิเจนโจมตีไขมันในเยื่อหุ้มเซลล์ ตัวรับโปรตีน เอนไซม์ และ DNA ซึ่งสามารถฆ่าไมโตคอนเดรียก่อนเวลาอันควร
บางที่จริงแล้วอนุมูลอิสระยังมีประโยชน์อีกด้วย ซึ่งจำเป็นสำหรับร่างกายในการควบคุมการทำงานของเซลล์ แต่ปัญหาเกิดขึ้นจากการก่อตัวของอนุมูลอิสระมากเกินไป น่าเสียดายที่นี่คือสาเหตุที่ประชากรส่วนใหญ่เกิดโรคส่วนใหญ่ โดยเฉพาะมะเร็ง มีสองวิธีในการแก้ปัญหานี้:
- เพิ่มสารต้านอนุมูลอิสระ
- ลดการผลิตอนุมูลอิสระจากไมโตคอนเดรีย
ในความคิดของฉัน หนึ่งในกลยุทธ์ที่มีประสิทธิภาพที่สุดในการลดอนุมูลอิสระในไมโตคอนเดรียคือการจำกัดปริมาณเชื้อเพลิงที่คุณใส่เข้าไปในร่างกาย สิ่งนี้ไม่เป็นที่ถกเถียงแต่อย่างใด เนื่องจากการจำกัดแคลอรี่แสดงให้เห็นประโยชน์ในการรักษาโรคหลายประการมาโดยตลอด นี่เป็นหนึ่งในเหตุผลที่การอดอาหารเป็นระยะมีประสิทธิภาพ เนื่องจากเป็นการจำกัดระยะเวลาในการบริโภคอาหาร ซึ่งจะช่วยลดปริมาณแคลอรี่ที่บริโภคโดยอัตโนมัติ
วิธีนี้จะได้ผลดีเป็นพิเศษหากคุณไม่ทานอาหารสองสามชั่วโมงก่อนนอน เพราะนี่เป็นสภาวะที่มีการเผาผลาญต่ำที่สุด
ทั้งหมดนี้อาจดูซับซ้อนเกินไปสำหรับผู้ที่ไม่ใช่ผู้เชี่ยวชาญ แต่สิ่งหนึ่งที่ต้องเข้าใจก็คือ เนื่องจากร่างกายใช้แคลอรี่น้อยที่สุดในระหว่างการนอนหลับ คุณจึงควรหลีกเลี่ยงการรับประทานอาหารก่อนนอน เพราะเชื้อเพลิงส่วนเกินในเวลานี้จะนำไปสู่การก่อตัวของปริมาณแคลอรี่ที่มากเกินไป อนุมูลอิสระที่ทำลายเนื้อเยื่อเร่งการแก่ชราและมีส่วนทำให้เกิดโรคเรื้อรัง
การอดอาหารช่วยให้การทำงานของไมโตคอนเดรียมีสุขภาพดีได้อย่างไร?
แพทริคยังตั้งข้อสังเกตอีกว่าส่วนหนึ่งของกลไกเบื้องหลังประสิทธิผลของการอดอาหารก็คือร่างกายถูกบังคับให้ได้รับพลังงานจากไขมันและไขมันสะสม ซึ่งหมายความว่าเซลล์จะถูกบังคับให้ใช้ไมโตคอนเดรีย
ไมโตคอนเดรียเป็นกลไกเดียวที่ร่างกายสามารถสร้างพลังงานจากไขมันได้ ดังนั้นการอดอาหารจึงช่วยกระตุ้นการทำงานของไมโตคอนเดรีย
นอกจากนี้เธอยังเชื่อว่ามันมีบทบาทอย่างมากในกลไกที่การอดอาหารเป็นระยะ ๆ และอาหารที่เป็นคีโตเจนิกฆ่าเซลล์มะเร็ง และอธิบายว่าทำไมยาที่กระตุ้นการทำงานของไมโตคอนเดรียบางชนิดจึงสามารถฆ่าเซลล์มะเร็งได้ อีกครั้ง นี่เป็นเพราะมีการผลิตออกซิเจนชนิดปฏิกิริยาเพิ่มขึ้น ความเสียหายที่ตัดสินผลลัพธ์ของสสาร ทำให้เกิดการตายของเซลล์มะเร็ง
โภชนาการของไมโตคอนเดรีย
จากมุมมองด้านโภชนาการ แพทริคเน้นย้ำถึงสารอาหารต่อไปนี้และปัจจัยร่วมที่สำคัญที่จำเป็นสำหรับการทำงานที่เหมาะสมของเอนไซม์ไมโตคอนเดรีย:
- Coenzyme Q10 หรือ ubiquinol (รูปแบบรีดิวซ์)
- L-carnitine ซึ่งขนส่งกรดไขมันเข้าสู่ไมโตคอนเดรีย
- D-ribose ซึ่งเป็นวัตถุดิบสำหรับโมเลกุล ATP
- แมกนีเซียม
- วิตามินบีทั้งหมด รวมถึงไรโบฟลาวิน ไทอามีน และบี 6
- กรดอัลฟ่าไลโปอิก (ALA)
ดังที่แพทริคตั้งข้อสังเกต:
“ฉันชอบได้รับสารอาหารรองมากที่สุดเท่าที่จะเป็นไปได้จากอาหารทั้งตัวด้วยเหตุผลหลายประการ ประการแรกพวกมันก่อตัวเป็นเส้นใยที่ซับซ้อนซึ่งเอื้อต่อการดูดซึม
นอกจากนี้ในกรณีนี้จะรับประกันอัตราส่วนที่ถูกต้อง คุณจะไม่สามารถรับมันได้มากมาย อัตราส่วนคือสิ่งที่คุณต้องการ ยังมีองค์ประกอบอื่นๆ ที่ยังรอการพิจารณาอยู่
คุณต้องระมัดระวังอย่างมากในการรับประทาน [อาหาร] ที่หลากหลาย และได้รับสารอาหารรองที่เหมาะสม ฉันคิดว่าการเสริมวิตามินบีรวมมีประโยชน์ด้วยเหตุนี้
ด้วยเหตุนี้ฉันจึงยอมรับพวกเขา อีกเหตุผลหนึ่งก็คือ เมื่อเราอายุมากขึ้น เราไม่สามารถดูดซึมวิตามินบีได้ง่ายอีกต่อไป สาเหตุหลักมาจากความแข็งของเยื่อหุ้มเซลล์ที่เพิ่มขึ้น สิ่งนี้จะเปลี่ยนวิธีการขนส่งวิตามินบีเข้าสู่เซลล์ ละลายน้ำได้จึงไม่สะสมอยู่ในไขมัน เป็นไปไม่ได้ที่จะถูกพวกมันวางยาพิษ ในกรณีที่รุนแรง คุณจะปัสสาวะมากขึ้นอีกเล็กน้อย แต่ฉันมั่นใจว่ามันมีประโยชน์มาก”
การออกกำลังกายสามารถช่วยให้ไมโตคอนเดรียยังเด็กได้
การออกกำลังกายยังส่งเสริมสุขภาพของไมโตคอนเดรียด้วยเพราะจะทำให้ไมโตคอนเดรียทำงานได้ ดังที่ได้กล่าวไว้ข้างต้น ผลข้างเคียงประการหนึ่งของกิจกรรมไมโตคอนเดรียที่เพิ่มขึ้นคือการสร้างสายพันธุ์ออกซิเจนที่เกิดปฏิกิริยา ซึ่งทำหน้าที่เป็นโมเลกุลส่งสัญญาณ
หน้าที่อย่างหนึ่งที่พวกมันส่งสัญญาณคือการก่อตัวของไมโตคอนเดรียมากขึ้น ดังนั้นเมื่อคุณออกกำลังกาย ร่างกายจะตอบสนองด้วยการสร้างไมโตคอนเดรียเพิ่มขึ้นเพื่อตอบสนองความต้องการพลังงานที่เพิ่มขึ้น
ความแก่ชราเป็นสิ่งที่หลีกเลี่ยงไม่ได้ แต่อายุทางชีวภาพของคุณอาจแตกต่างอย่างมากจากอายุตามลำดับเวลา และไมโตคอนเดรียก็มีความคล้ายคลึงกับอายุทางชีวภาพเป็นอย่างมาก แพทริคอ้างอิงงานวิจัยล่าสุดที่แสดงให้เห็นว่าผู้คนสามารถสูงวัยตามหลักชีววิทยาได้อย่างไร มากในจังหวะที่ต่างกัน
นักวิจัยตรวจวัดตัวบ่งชี้ทางชีวภาพที่แตกต่างกันมากกว่าหนึ่งโหล เช่น ความยาวของเทโลเมียร์ ความเสียหายของดีเอ็นเอ คอเลสเตอรอลชนิดไม่ดี เมแทบอลิซึมของกลูโคส และความไวต่ออินซูลิน ณ จุดสามจุดในชีวิตของผู้คน ได้แก่ อายุ 22, 32 และ 38 ปี
“เราพบว่าคนที่มีอายุ 38 ปีอาจมีหน้าตาที่ดูอ่อนกว่าวัยหรือแก่กว่านั้นได้ถึง 10 ปี โดยพิจารณาจากเครื่องหมายทางชีวภาพ แม้จะอายุเท่ากัน ความชราทางชีวภาพก็เกิดขึ้นในอัตราที่ต่างกันโดยสิ้นเชิง
สิ่งที่น่าสนใจคือเมื่อถ่ายภาพคนเหล่านี้และแสดงภาพถ่ายของพวกเขาให้ผู้คนที่เดินผ่านไปมาเห็น และขอให้เดาอายุตามลำดับเวลาของบุคคลที่ปรากฎ ผู้คนจะเดาอายุทางชีววิทยา ไม่ใช่อายุตามลำดับเวลา”
ดังนั้น ไม่ว่าคุณจะอายุเท่าใด รูปร่างหน้าตาของคุณก็สอดคล้องกับตัวบ่งชี้ทางชีวภาพของคุณ ซึ่งส่วนใหญ่จะถูกกำหนดโดยสุขภาพของไมโตคอนเดรีย ดังนั้นในขณะที่ไม่สามารถหลีกเลี่ยงความชราได้ แต่คุณสามารถควบคุมอายุได้มาก และนั่นก็มีพลังมาก และหนึ่งในปัจจัยสำคัญคือการรักษาไมโตคอนเดรียให้อยู่ในสภาพทำงานได้ดี
ตามที่แพทริคกล่าวไว้ “วัยเยาว์” ไม่ใช่อายุตามลำดับเวลามากนัก แต่ขึ้นอยู่กับอายุที่คุณรู้สึกและร่างกายของคุณทำงานได้ดีเพียงใด:
“ฉันต้องการทราบวิธีเพิ่มประสิทธิภาพการทำงานของจิตใจและสมรรถภาพทางกีฬาของฉัน ฉันต้องการยืดอายุความเยาว์วัยของฉัน ฉันอยากจะมีอายุถึง 90 ปี และเมื่อฉันทำได้ ฉันอยากจะเล่นเซิร์ฟในซานดิเอโกแบบเดียวกับตอนอายุ 20 ฉันหวังว่าฉันจะไม่หายไปเร็วเหมือนบางคน ฉันชอบที่จะชะลอความเสื่อมถอยนี้และยืดอายุความเยาว์วัยของฉันให้นานที่สุด เพื่อที่ฉันจะได้มีความสุขกับชีวิตมากที่สุด”
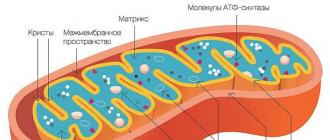
โครงสร้าง. อุปกรณ์พื้นผิวของไมโตคอนเดรียประกอบด้วยเยื่อหุ้มสองอัน - ด้านนอกและด้านใน เยื่อหุ้มชั้นนอกเรียบเนียน แยกไมโตคอนเดรียออกจากไฮยาโลพลาสซึม ข้างใต้เป็นรอยพับ เยื่อหุ้มชั้นใน,แบบฟอร์มไหน คริสตี้(สันเขา). ทั้งสองด้านของคริสเต มีลำตัวคล้ายเห็ดเล็กๆ เรียกว่า ออกซิโซม หรือ ATP-โซมามิประกอบด้วยเอนไซม์ที่เกี่ยวข้องกับออกซิเดชั่นฟอสโฟรีเลชั่น (การเติมฟอสเฟตที่ตกค้างใน ADP เพื่อสร้าง ATP) จำนวนคริสเตในไมโตคอนเดรียสัมพันธ์กับความต้องการพลังงานของเซลล์ โดยเฉพาะอย่างยิ่งในเซลล์กล้ามเนื้อ ไมโตคอนเดรียมีคริสเตจำนวนมาก ด้วยการทำงานของเซลล์ที่เพิ่มขึ้น ไมโตคอนเดรียจะกลายเป็นรูปไข่หรือยาวขึ้น และจำนวนคริสเตก็เพิ่มขึ้น
ไมโตคอนเดรียมีจีโนมของตัวเอง ไรโบโซมประเภท 70S แตกต่างจากไรโบโซมของไซโตพลาสซึม DNA ของไมโตคอนเดรียส่วนใหญ่มีรูปแบบไซคลิก (พลาสมิด) เข้ารหัส RNA ทั้งสามประเภทของตัวเอง และให้ข้อมูลสำหรับการสังเคราะห์โปรตีนไมโตคอนเดรียบางชนิด (ประมาณ 9%) ดังนั้นไมโตคอนเดรียจึงถือเป็นออร์แกเนลล์กึ่งอิสระได้ ไมโตคอนเดรียเป็นออร์แกเนลล์ที่จำลองตัวเองได้ (สามารถสืบพันธุ์ได้) การต่ออายุของไมโตคอนเดรียเกิดขึ้นตลอดวัฏจักรของเซลล์ ตัวอย่างเช่นในเซลล์ตับจะถูกแทนที่ด้วยเซลล์ใหม่หลังจากผ่านไปเกือบ 10 วัน วิธีที่เป็นไปได้มากที่สุดในการสร้างไมโตคอนเดรียนั้นถือเป็นการแบ่งตัว: การหดตัวปรากฏขึ้นตรงกลางไมโตคอนเดรียหรือมีกะบังปรากฏขึ้น หลังจากนั้นออร์แกเนลล์ก็แยกออกเป็นไมโตคอนเดรียใหม่สองตัว ไมโตคอนเดรียถูกสร้างขึ้นด้วยโพรมิโตคอนเดรีย - วัตถุทรงกลมที่มีเส้นผ่านศูนย์กลางสูงถึง 50 นาโนเมตรพร้อมเมมเบรนสองชั้น
ฟังก์ชั่น . ไมโตคอนเดรียเกี่ยวข้องกับกระบวนการพลังงานของเซลล์ ไมโตคอนเดรียประกอบด้วยเอนไซม์ที่เกี่ยวข้องกับการผลิตพลังงานและการหายใจของเซลล์ กล่าวอีกนัยหนึ่ง ไมโตคอนเดรียเป็นโรงงานขนาดเล็กทางชีวเคมีชนิดหนึ่งที่แปลงพลังงานของสารประกอบอินทรีย์ให้เป็นพลังงานที่ใช้ของ ATP ในไมโตคอนเดรีย กระบวนการพลังงานเริ่มต้นในเมทริกซ์ ซึ่งการสลายของกรดไพรูวิกเกิดขึ้นในวงจรเครบส์ ในระหว่างกระบวนการนี้ อะตอมของไฮโดรเจนจะถูกปล่อยและขนส่งโดยระบบทางเดินหายใจ พลังงานที่ปล่อยออกมาในกรณีนี้ถูกใช้ในหลายส่วนของห่วงโซ่การหายใจเพื่อทำปฏิกิริยาฟอสโฟรีเลชั่น - การสังเคราะห์ ATP นั่นคือการเติมกลุ่มฟอสเฟตใน ADP สิ่งนี้เกิดขึ้นที่เยื่อหุ้มชั้นในของไมโตคอนเดรีย ดังนั้น, ฟังก์ชั่นพลังงานไมโตคอนเดรียรวมเข้ากับ: ก) การออกซิเดชั่นของสารประกอบอินทรีย์ที่เกิดขึ้นในเมทริกซ์เนื่องจากการเรียกไมโตคอนเดรีย ศูนย์กลางการหายใจของเซลล์ b) การสังเคราะห์ ATP ดำเนินการกับคริสเตเนื่องจากการเรียกไมโตคอนเดรีย สถานีพลังงานของเซลล์นอกจากนี้ไมโตคอนเดรียยังมีส่วนร่วมในการควบคุมการเผาผลาญของน้ำ, การสะสมของแคลเซียมไอออน, การผลิตสารตั้งต้นของฮอร์โมนสเตียรอยด์, เมแทบอลิซึม (เช่นไมโตคอนเดรียในเซลล์ตับมีเอนไซม์ที่ช่วยให้พวกมันต่อต้านแอมโมเนีย) และอื่น ๆ
ชีววิทยา + โรคไมโตคอนเดรียเป็นกลุ่มของโรคทางพันธุกรรมที่เกี่ยวข้องกับข้อบกพร่องของไมโตคอนเดรียที่นำไปสู่การหายใจของเซลล์บกพร่อง พวกมันจะถูกส่งผ่านสายตัวเมียไปยังเด็กทั้งสองเพศ เนื่องจากไข่มีไซโตพลาสซึมในปริมาณที่มากกว่าและด้วยเหตุนี้จึงส่งผ่านไมโตคอนเดรียจำนวนมากไปยังลูกหลาน DNA ไมโตคอนเดรียซึ่งแตกต่างจาก DNA นิวเคลียร์ไม่ได้รับการปกป้องโดยโปรตีนฮิสโตน และกลไกการซ่อมแซมที่สืบทอดมาจากแบคทีเรียของบรรพบุรุษนั้นไม่สมบูรณ์ ดังนั้นการกลายพันธุ์จึงสะสมใน DNA ของไมโตคอนเดรียเร็วกว่า DNA นิวเคลียร์ถึง 10-20 เท่า ซึ่งนำไปสู่โรคของไมโตคอนเดรีย ในการแพทย์แผนปัจจุบัน มีประมาณ 50 ชนิดที่ทราบกันดีอยู่แล้ว ตัวอย่างเช่น กลุ่มอาการอ่อนเพลียเรื้อรัง ไมเกรน กลุ่มอาการบาร์ธ กลุ่มอาการเพียร์สัน และอื่นๆ อีกมากมาย
ไมโตคอนเดรียเป็นออร์แกเนลล์ที่จับกับเมมเบรนด้วยกล้องจุลทรรศน์ที่ให้พลังงานแก่เซลล์ ดังนั้นจึงเรียกว่าสถานีพลังงาน (แบตเตอรี่) ของเซลล์
ไมโตคอนเดรียไม่อยู่ในเซลล์ของสิ่งมีชีวิตธรรมดา แบคทีเรีย และเอนทามีบา ซึ่งมีชีวิตอยู่โดยไม่ต้องใช้ออกซิเจน สาหร่ายสีเขียวบางชนิด ทริปาโนโซมมีไมโตคอนเดรียขนาดใหญ่ 1 ตัว และเซลล์ของกล้ามเนื้อหัวใจและสมองมีออร์แกเนลล์เหล่านี้ตั้งแต่ 100 ถึง 1,000 ตัว
คุณสมบัติโครงสร้าง
ไมโตคอนเดรียเป็นออร์แกเนลล์ที่มีเยื่อหุ้มสองชั้น มีเยื่อหุ้มชั้นนอกและชั้นใน มีช่องว่างระหว่างเยื่อหุ้มเซลล์ และเมทริกซ์
เยื่อหุ้มชั้นนอก- มีความเรียบ ไม่มีรอยพับ และแยกสิ่งที่อยู่ภายในออกจากไซโตพลาสซึม ความกว้างของมันคือ 7 นาโนเมตรและมีไขมันและโปรตีน มีบทบาทสำคัญในการเล่นโดย porin ซึ่งเป็นโปรตีนที่สร้างช่องทางในเยื่อหุ้มชั้นนอก พวกมันให้การแลกเปลี่ยนไอออนและโมเลกุล
พื้นที่ระหว่างเมมเบรน- ขนาดของช่องว่างระหว่างเมมเบรนคือประมาณ 20 นาโนเมตร สารที่บรรจุนั้นมีองค์ประกอบคล้ายคลึงกับไซโตพลาสซึมยกเว้นโมเลกุลขนาดใหญ่ที่สามารถแทรกซึมได้ที่นี่ผ่านการขนส่งแบบแอคทีฟเท่านั้น

เยื่อหุ้มชั้นใน- ประกอบด้วยโปรตีนเป็นส่วนใหญ่ โดยมีเพียง 1 ใน 3 เท่านั้นที่ถูกจัดสรรให้กับสารไขมัน โปรตีนจำนวนมากเป็นโปรตีนในการขนส่งเนื่องจากเยื่อหุ้มชั้นในไม่มีรูขุมขนที่ผ่านได้อย่างอิสระ มันก่อให้เกิดผลพลอยได้หลายอย่าง - คริสเตซึ่งดูเหมือนสันเขาแบน ออกซิเดชันของสารประกอบอินทรีย์กับ CO 2 ในไมโตคอนเดรียเกิดขึ้นบนเยื่อหุ้มของคริสเต กระบวนการนี้ขึ้นอยู่กับออกซิเจนและดำเนินการภายใต้การกระทำของ ATP synthetase พลังงานที่ปล่อยออกมาจะถูกเก็บไว้ในรูปของโมเลกุล ATP และนำไปใช้ตามความจำเป็น
เมทริกซ์– สภาพแวดล้อมภายในของไมโตคอนเดรียมีโครงสร้างเป็นเนื้อเดียวกันเป็นเม็ดละเอียด ในกล้องจุลทรรศน์อิเล็กตรอน คุณสามารถมองเห็นแกรนูลและเส้นใยในลูกบอลที่วางอย่างอิสระระหว่างคริสเต เมทริกซ์มีระบบสังเคราะห์โปรตีนกึ่งอัตโนมัติ - DNA, RNA ทุกประเภทและไรโบโซมอยู่ที่นี่ แต่ถึงกระนั้นโปรตีนส่วนใหญ่ได้มาจากนิวเคลียส ซึ่งเป็นเหตุให้ไมโตคอนเดรียถูกเรียกว่าออร์แกเนลล์กึ่งอิสระ
ตำแหน่งและการแบ่งเซลล์
ฮอนดริโอมคือกลุ่มไมโตคอนเดรียที่รวมตัวกันอยู่ในเซลล์เดียว พวกมันอยู่ในไซโตพลาสซึมที่แตกต่างกันซึ่งขึ้นอยู่กับความเชี่ยวชาญของเซลล์ ตำแหน่งในไซโตพลาสซึมยังขึ้นอยู่กับออร์แกเนลล์โดยรอบและการรวมเข้าด้วยกัน ในเซลล์พืชพวกมันครอบครองบริเวณรอบนอกเนื่องจากแวคิวโอลส่วนกลางผลักไมโตคอนเดรียไปยังเยื่อหุ้มเซลล์ ในเซลล์เยื่อบุผิวไตเมมเบรนจะยื่นออกมาซึ่งระหว่างนั้นจะมีไมโตคอนเดรีย
ในเซลล์ต้นกำเนิดซึ่งออร์แกเนลล์ทุกตัวใช้พลังงานเท่ากัน ไมโตคอนเดรียจะถูกกระจายแบบสุ่ม ในเซลล์เฉพาะทาง พวกมันจะกระจุกตัวอยู่ในพื้นที่ที่ใช้พลังงานมากที่สุดเป็นหลัก ตัวอย่างเช่นในกล้ามเนื้อโครงร่างพวกมันจะอยู่ใกล้กับไมโอไฟบริล ในตัวอสุจิ พวกมันจะปกคลุมแกนของแฟลเจลลัมเป็นเกลียว เนื่องจากจำเป็นต้องใช้พลังงานจำนวนมากในการทำให้ตัวอสุจิเคลื่อนไหวและเคลื่อนไหว โปรโตซัวที่เคลื่อนที่โดยใช้ซีเลียยังมีไมโตคอนเดรียจำนวนมากที่ฐาน
แผนก- ไมโตคอนเดรียมีความสามารถในการสืบพันธุ์แบบอิสระและมีจีโนมของตัวเอง Organelles แบ่งตามการหดตัวหรือผนังกั้น การก่อตัวของไมโตคอนเดรียใหม่ในเซลล์ต่างๆ จะมีความถี่แตกต่างกัน เช่น ในเนื้อเยื่อตับ ไมโตคอนเดรียจะถูกแทนที่ทุกๆ 10 วัน
หน้าที่ในเซลล์
- หน้าที่หลักของไมโตคอนเดรียคือการก่อตัวของโมเลกุลเอทีพี
- การสะสมของแคลเซียมไอออน
- การมีส่วนร่วมแลกเปลี่ยนน้ำ
- การสังเคราะห์สารตั้งต้นของฮอร์โมนสเตียรอยด์
อณูชีววิทยาเป็นวิทยาศาสตร์ที่ศึกษาบทบาทของไมโตคอนเดรียในการเผาผลาญ พวกเขายังเปลี่ยนไพรูเวตเป็นอะซิติลโคเอ็นไซม์เอและเบต้าออกซิเดชันของกรดไขมัน
| ตาราง: โครงสร้างและหน้าที่ของไมโตคอนเดรีย (สั้น ๆ ) | ||
|---|---|---|
| องค์ประกอบโครงสร้าง | โครงสร้าง | ฟังก์ชั่น |
| เยื่อหุ้มชั้นนอก | เปลือกเรียบทำจากไขมันและโปรตีน | แยกเนื้อหาภายในออกจากไซโตพลาสซึม |
| พื้นที่ระหว่างเมมเบรน | มีไฮโดรเจนไอออน โปรตีน ไมโครโมเลกุล | สร้างการไล่ระดับโปรตอน |
| เยื่อหุ้มชั้นใน | ยื่นออกมาในรูปแบบ - คริสเต มีระบบขนส่งโปรตีน | การถ่ายโอนโมเลกุลขนาดใหญ่ การคงสภาพการไล่ระดับโปรตอน |
| เมทริกซ์ | ตำแหน่งของเอนไซม์รอบเครบส์, DNA, RNA, ไรโบโซม | ออกซิเดชันแบบแอโรบิกด้วยการปลดปล่อยพลังงาน การแปลงไพรูเวตเป็นอะเซทิลโคเอ็นไซม์เอ |
| ไรโบโซม | รวมสองหน่วยย่อยเข้าด้วยกัน | การสังเคราะห์โปรตีน |
ความคล้ายคลึงกันระหว่างไมโตคอนเดรียและคลอโรพลาสต์
คุณสมบัติทั่วไปของไมโตคอนเดรียและคลอโรพลาสต์มีสาเหตุหลักมาจากการมีเมมเบรนสองชั้น
สัญญาณของความคล้ายคลึงกันยังรวมถึงความสามารถในการสังเคราะห์โปรตีนอย่างอิสระ ออร์แกเนลล์เหล่านี้มี DNA, RNA และไรโบโซมของตัวเอง
ทั้งไมโตคอนเดรียและคลอโรพลาสต์สามารถแบ่งตัวได้ด้วยการหดตัว
พวกมันยังรวมกันเป็นหนึ่งด้วยความสามารถในการผลิตพลังงาน ไมโตคอนเดรียมีความเชี่ยวชาญมากกว่าในฟังก์ชันนี้ แต่คลอโรพลาสต์ยังผลิตโมเลกุล ATP ในระหว่างกระบวนการสังเคราะห์แสง ดังนั้นเซลล์พืชจึงมีไมโตคอนเดรียน้อยกว่าเซลล์สัตว์ เนื่องจากคลอโรพลาสต์ทำหน้าที่บางส่วนให้กับพวกมัน
ให้เราอธิบายความเหมือนและความแตกต่างโดยย่อ:
- พวกมันเป็นออร์แกเนลล์ที่มีเยื่อหุ้มสองชั้น
- เยื่อหุ้มชั้นในก่อให้เกิดส่วนที่ยื่นออกมา: คริสเตเป็นลักษณะของไมโตคอนเดรียและไทลาคอยด์เป็นลักษณะของคลอโรพลาสต์
- มีจีโนมของตัวเอง
- สามารถสังเคราะห์โปรตีนและพลังงานได้
ออร์แกเนลล์เหล่านี้มีหน้าที่ต่างกัน: ไมโตคอนเดรียมีไว้สำหรับการสังเคราะห์พลังงาน การหายใจของเซลล์เกิดขึ้นที่นี่ เซลล์พืชต้องการคลอโรพลาสต์สำหรับการสังเคราะห์ด้วยแสง
ไมโตคอนเดรีย (จากภาษากรีก μίτος (mitos) - ด้าย และ χονδρίον (chondrion) - เม็ดเล็ก) เป็นออร์แกเนลล์สองเยื่อหุ้มเซลล์ที่มีสารพันธุกรรมของตัวเอง ซึ่งก็คือไมโตคอนเดรีย พบเป็นโครงสร้างเซลล์ทรงกลมหรือท่อในยูคาริโอตเกือบทั้งหมด แต่ไม่ใช่ในโปรคาริโอต
ไมโตคอนเดรียเป็นออร์แกเนลล์ที่สร้างโมเลกุลอะดีโนซีน ไตรฟอสเฟต พลังงานสูงขึ้นมาใหม่ผ่านระบบทางเดินหายใจ นอกเหนือจากออกซิเดชันฟอสโฟรีเลชันแล้ว ยังทำหน้าที่สำคัญอื่นๆ เช่น มีส่วนร่วมในการก่อตัวของกลุ่มเหล็กและกำมะถัน- โครงสร้างและหน้าที่ของออร์แกเนลล์ดังกล่าวมีรายละเอียดดังนี้
ติดต่อกับ
ข้อมูลทั่วไป
มีไมโตคอนเดรียจำนวนมากโดยเฉพาะในพื้นที่ที่มีการใช้พลังงานสูง ซึ่งรวมถึงกล้ามเนื้อ เส้นประสาท เซลล์รับความรู้สึก และโอโอไซต์ ในโครงสร้างเซลล์ของกล้ามเนื้อหัวใจ ปริมาตรของออร์แกเนลล์เหล่านี้สูงถึง 36% มีเส้นผ่านศูนย์กลางประมาณ 0.5-1.5 ไมครอน และมีรูปร่างหลากหลายตั้งแต่ทรงกลมไปจนถึงเกลียวที่ซับซ้อน จำนวนของพวกเขาจะถูกปรับโดยคำนึงถึงความต้องการพลังงานของเซลล์
เซลล์ยูคาริโอตที่สูญเสียไมโตคอนเดรีย ไม่สามารถกู้คืนได้- นอกจากนี้ยังมียูคาริโอตที่ไม่มีพวกมัน เช่น โปรโตซัวบางชนิด จำนวนออร์แกเนลล์เหล่านี้ต่อหน่วยเซลล์มักจะอยู่ระหว่าง 1,000 ถึง 2,000 โดยมีเศษส่วนปริมาตร 25% แต่ค่าเหล่านี้อาจแตกต่างกันอย่างมากขึ้นอยู่กับชนิดของโครงสร้างเซลล์และสิ่งมีชีวิต มีประมาณสี่ถึงห้าตัวในเซลล์อสุจิที่โตเต็มที่ และหลายแสนตัวในไข่ที่โตเต็มที่
ไมโตคอนเดรียถูกส่งผ่านพลาสมาของไข่จากแม่เท่านั้น ซึ่งเป็นเหตุผลในการศึกษาสายเลือดของมารดา ปัจจุบันเป็นที่ยอมรับแล้วว่าออร์แกเนลล์ตัวผู้บางตัวถูกนำเข้าสู่พลาสมาของไข่ที่ปฏิสนธิ (ไซโกต) ผ่านสเปิร์มด้วย พวกเขาคงจะได้รับการแก้ไขอย่างรวดเร็ว อย่างไรก็ตาม มีหลายกรณีที่แพทย์สามารถพิสูจน์ได้ว่าไมโตคอนเดรียของเด็กนั้นมาจากสายเลือดบิดา โรคที่เกิดจากการกลายพันธุ์ของยีนไมโตคอนเดรียนั้นถ่ายทอดมาจากแม่เท่านั้น
น่าสนใจ!คำศัพท์ทางวิทยาศาสตร์ยอดนิยม "โรงไฟฟ้าของเซลล์" ได้รับการประกาศเกียรติคุณในปี 1957 โดย Philip Sikiewitz
แผนภาพโครงสร้างไมโตคอนเดรีย
ให้เราพิจารณาลักษณะโครงสร้างของโครงสร้างที่สำคัญเหล่านี้ พวกมันถูกสร้างขึ้นจากการรวมกันขององค์ประกอบหลายอย่าง เปลือกของออร์แกเนลล์เหล่านี้ประกอบด้วยเยื่อหุ้มชั้นนอกและชั้นใน ในทางกลับกัน ประกอบด้วยชั้นสองของฟอสโฟไลปิดและโปรตีน เปลือกหอยทั้งสองมีคุณสมบัติต่างกัน ระหว่างนั้นมีช่องที่แตกต่างกันห้าช่อง: เยื่อหุ้มชั้นนอก, ช่องว่างระหว่างเยื่อหุ้มเซลล์ (ช่องว่างระหว่างเยื่อหุ้มทั้งสอง), เยื่อหุ้มชั้นใน, คริสตาและเมทริกซ์ (ช่องว่างภายในเยื่อหุ้มชั้นใน) โดยทั่วไป - โครงสร้างภายในของออร์แกเนลล์ .
ในภาพประกอบในตำราเรียน ไมโตคอนเดรียส่วนใหญ่จะดูเหมือนออร์แกเนลล์รูปถั่วที่แยกจากกัน จริงเหรอ? ไม่ พวกมันก่อตัว เครือข่ายไมโตคอนเดรียแบบท่อซึ่งสามารถทะลุผ่านและเปลี่ยนหน่วยเซลลูลาร์ทั้งหมดได้ ไมโตคอนเดรียในเซลล์สามารถรวม (โดยการหลอมรวม) และการแบ่งตัวใหม่ (ฟิชชัน)
บันทึก!ในยีสต์ การหลอมรวมไมโตคอนเดรียประมาณสองครั้งเกิดขึ้นในหนึ่งนาที ดังนั้นจึงเป็นไปไม่ได้ที่จะระบุจำนวนไมโตคอนเดรียในเซลล์ในปัจจุบันได้อย่างแม่นยำ
เยื่อหุ้มชั้นนอก
 เปลือกนอกล้อมรอบออร์แกเนลล์ทั้งหมดและมีช่องของโปรตีนเชิงซ้อนที่ช่วยให้เกิดการแลกเปลี่ยนโมเลกุลและไอออนระหว่างไมโตคอนเดรียและไซโตซอล โมเลกุลขนาดใหญ่ ไม่สามารถผ่านเมมเบรนได้.
เปลือกนอกล้อมรอบออร์แกเนลล์ทั้งหมดและมีช่องของโปรตีนเชิงซ้อนที่ช่วยให้เกิดการแลกเปลี่ยนโมเลกุลและไอออนระหว่างไมโตคอนเดรียและไซโตซอล โมเลกุลขนาดใหญ่ ไม่สามารถผ่านเมมเบรนได้.
ชั้นนอกซึ่งทอดยาวไปทั่วทั้งออร์แกเนลล์และไม่พับ มีอัตราส่วนฟอสโฟไลปิดต่อน้ำหนักโปรตีน 1:1 จึงคล้ายกับพลาสมาเมมเบรนยูคาริโอต ประกอบด้วยโปรตีนและพอรินหลายชนิด Porins สร้างช่องทางที่ช่วยให้โมเลกุลแพร่กระจายอย่างอิสระโดยมีมวลมากถึง 5,000 ดาลตันผ่านเมมเบรน โปรตีนที่มีขนาดใหญ่กว่าสามารถบุกรุกได้เมื่อลำดับสัญญาณที่ปลาย N จับกับหน่วยย่อยขนาดใหญ่ของโปรตีนทรานส์ล็อกเซส จากนั้นพวกมันจะเคลื่อนตัวไปตามเปลือกเมมเบรนอย่างแข็งขัน
หากรอยแตกร้าวเกิดขึ้นในเยื่อหุ้มชั้นนอก โปรตีนจากช่องว่างระหว่างเมมเบรนสามารถหลุดออกไปในไซโตโซลได้ อาจทำให้เซลล์ตายได้- เมมเบรนด้านนอกสามารถหลอมรวมกับเยื่อเอนโดพลาสมิกเรติคูลัม จากนั้นจึงสร้างโครงสร้างที่เรียกว่า MAM (ER ที่เกี่ยวข้องกับไมโตคอนเดรีย) เป็นสิ่งสำคัญสำหรับการส่งสัญญาณระหว่าง ER และไมโตคอนเดรีย ซึ่งจำเป็นสำหรับการขนส่งด้วย
พื้นที่ระหว่างเมมเบรน
บริเวณที่เป็นช่องว่างระหว่างเยื่อหุ้มชั้นนอกและชั้นใน เนื่องจากโมเลกุลภายนอกช่วยให้โมเลกุลขนาดเล็กสามารถแทรกซึมได้อย่างอิสระ ความเข้มข้นของพวกมัน เช่น ไอออนและน้ำตาล ในพื้นที่ระหว่างเมมเบรนจึงเหมือนกับความเข้มข้นในไซโตโซล อย่างไรก็ตาม โปรตีนขนาดใหญ่จำเป็นต้องมีการถ่ายทอดลำดับสัญญาณที่เฉพาะเจาะจง ดังนั้นองค์ประกอบของโปรตีนจึงแตกต่างกันระหว่างช่องว่างระหว่างเมมเบรนและไซโตซอล ดังนั้นโปรตีนที่ยังคงอยู่ในช่องว่างระหว่างเมมเบรนคือไซโตโครม
เยื่อหุ้มชั้นใน
เยื่อหุ้มไมโตคอนเดรียชั้นในประกอบด้วยโปรตีนที่มีหน้าที่สี่ประเภท:
- โปรตีน - ทำปฏิกิริยาออกซิเดชั่นของห่วงโซ่ทางเดินหายใจ
- อะดีโนซีน ไตรฟอสเฟต ซินเทส ซึ่งสร้าง ATP ในเมทริกซ์
- โปรตีนขนส่งจำเพาะที่ควบคุมการผ่านของสารระหว่างเมทริกซ์และไซโตพลาสซึม
- ระบบนำเข้าโปรตีน
โดยเฉพาะอย่างยิ่งภายในมีฟอสโฟไลปิดคู่ คาร์ดิโอลิพิน แทนที่ด้วยกรดไขมันสี่ตัว Cardiolipin มักพบในเยื่อหุ้มไมโตคอนเดรียและเยื่อหุ้มพลาสมาของแบคทีเรีย มีอยู่ในร่างกายมนุษย์เป็นส่วนใหญ่ ในบริเวณที่มีกิจกรรมการเผาผลาญสูงหรือมีกิจกรรมที่มีพลังงานสูง เช่น กล้ามเนื้อหัวใจหดตัวในกล้ามเนื้อหัวใจ
ความสนใจ!เยื่อหุ้มชั้นในประกอบด้วยโพลีเปปไทด์ที่แตกต่างกันมากกว่า 150 ชนิด หรือประมาณ 1/8 ของโปรตีนไมโตคอนเดรียทั้งหมด ส่งผลให้ความเข้มข้นของไขมันต่ำกว่าชั้นนอกและความสามารถในการซึมผ่านของไขมันต่ำกว่า
 เมื่อแบ่งออกเป็นคริสเตจำนวนมาก พวกมันจะขยายบริเวณด้านนอกของเยื่อหุ้มไมโตคอนเดรียชั้นใน และเพิ่มความสามารถในการผลิต ATP
เมื่อแบ่งออกเป็นคริสเตจำนวนมาก พวกมันจะขยายบริเวณด้านนอกของเยื่อหุ้มไมโตคอนเดรียชั้นใน และเพิ่มความสามารถในการผลิต ATP
ในไมโตคอนเดรียในตับทั่วไป ตัวอย่างเช่น บริเวณด้านนอก โดยเฉพาะคริสเต มีขนาดประมาณห้าเท่าของพื้นที่เยื่อหุ้มชั้นนอก สถานีพลังงานของเซลล์ที่มีความต้องการ ATP สูงกว่า เช่น เซลล์กล้ามเนื้อมีคริสเตมากขึ้นมากกว่าไมโตคอนเดรียในตับทั่วไป
เยื่อหุ้มชั้นในล้อมรอบเมทริกซ์ ซึ่งเป็นของเหลวชั้นในของไมโตคอนเดรีย มันสอดคล้องกับไซโตโซลของแบคทีเรียและมี DNA ของไมโตคอนเดรีย เอนไซม์ซิเตรตและไรโบโซมไมโตคอนเดรียของพวกมันเอง ซึ่งแตกต่างจากไรโบโซมในไซโตโซล (แต่จากแบคทีเรียด้วย) พื้นที่ระหว่างเมมเบรนประกอบด้วยเอนไซม์ที่สามารถฟอสโฟรีเลทนิวคลีโอไทด์โดยการบริโภค ATP
ฟังก์ชั่น
- เส้นทางการย่อยสลายที่สำคัญ: วงจรซิเตรต ซึ่งมีการนำไพรูเวตจากไซโตโซลเข้าสู่เมทริกซ์ จากนั้นไพรูเวตจะถูกดีคาร์บอกซิเลตโดยไพรูเวตดีไฮโดรจีเนสไปเป็นอะซีติลโคเอ็นไซม์เอ อีกแหล่งหนึ่งของอะซิติลโคเอ็นไซม์เอคือการย่อยสลายกรดไขมัน (β-ออกซิเดชัน) ซึ่งเกิดขึ้นในเซลล์สัตว์ในไมโตคอนเดรีย แต่ในเซลล์พืชเฉพาะในไกลออกซีโซมและเปอร์รอกซิโซม เพื่อจุดประสงค์นี้ อะซิล-โคเอ็นไซม์ A จะถูกถ่ายโอนจากไซโตโซลโดยจับกับคาร์นิทีนผ่านเยื่อหุ้มไมโตคอนเดรียชั้นใน และแปลงเป็นอะซิติล-โคเอ็นไซม์ A จากนี้ ค่ารีดิวซ์ส่วนใหญ่ที่เทียบเท่ากันในวงจรเครบส์ (หรือเรียกอีกอย่างว่าวงจรเครบส์หรือ วงจรกรดไตรคาร์บอกซิลิก) ซึ่งจะถูกแปลงเป็น ATP ในสายออกซิเดชัน
- โซ่ออกซิเดชั่น การไล่ระดับเคมีไฟฟ้าถูกสร้างขึ้นระหว่างสเปซอินเทอร์เมมเบรนและเมทริกซ์ไมโตคอนเดรีย ซึ่งทำหน้าที่สร้าง ATP โดยใช้ ATP synthase ผ่านกระบวนการถ่ายโอนอิเล็กตรอนและการสะสมโปรตอน จะได้อิเล็กตรอนและโปรตอนที่จำเป็นในการสร้างการไล่ระดับสี โดยการย่อยสลายสารอาหารด้วยปฏิกิริยาออกซิเดชั่น(เช่นกลูโคส) ที่ร่างกายดูดซึมได้ ไกลโคไลซิสเริ่มแรกเกิดขึ้นในไซโตพลาสซึม
- อะพอพโทซิส (การตายของเซลล์ที่ตั้งโปรแกรมไว้)
- การเก็บแคลเซียม: ไมโตคอนเดรียจะเข้าไปแทรกแซงสภาวะสมดุลของเซลล์ด้วยความสามารถในการดูดซับไอออนแคลเซียมแล้วปล่อยออกมา
- การสังเคราะห์กลุ่มเหล็ก-ซัลเฟอร์จำเป็นต้องใช้เอนไซม์หลายชนิดในห่วงโซ่ทางเดินหายใจ ฟังก์ชันนี้ถือเป็นฟังก์ชันสำคัญของไมโตคอนเดรีย กล่าวคือ นี่คือเหตุผลว่าทำไมเซลล์เกือบทั้งหมดต้องอาศัยสถานีพลังงานเพื่อความอยู่รอด
![]()
เมทริกซ์
นี่คือช่องว่างที่รวมอยู่ในเยื่อหุ้มไมโตคอนเดรียชั้นใน มีโปรตีนประมาณสองในสามของทั้งหมด มีบทบาทสำคัญในการผลิต ATP ผ่านทาง ATP synthase ซึ่งรวมอยู่ในเยื่อหุ้มชั้นใน ประกอบด้วยเอนไซม์หลายร้อยชนิดที่มีความเข้มข้นสูง (ส่วนใหญ่เกี่ยวข้องกับการย่อยสลายกรดไขมันและไพรูเวต), ไรโบโซมที่จำเพาะต่อไมโตคอนเดรีย, Messenger RNA และสำเนา DNA ของจีโนมไมโตคอนเดรียหลายชุด
ออร์แกเนลล์เหล่านี้มีจีโนมของตัวเอง เช่นเดียวกับอุปกรณ์เอนไซม์ที่จำเป็นสำหรับ ดำเนินการสังเคราะห์โปรตีนด้วยตัวมันเอง.
ไมโตคอนเดรีย ไมโตคอนเดรียคืออะไร พร้อมหน้าที่ของมัน
โครงสร้างและการทำงานของไมโตคอนเดรีย
บทสรุป
ดังนั้นไมโตคอนเดรียจึงถูกเรียกว่าโรงไฟฟ้าพลังงานเซลล์ที่ผลิตพลังงานและครองตำแหน่งผู้นำในชีวิตและการอยู่รอดของเซลล์แต่ละเซลล์โดยเฉพาะและสิ่งมีชีวิตโดยทั่วไป ไมโตคอนเดรียเป็นส่วนสำคัญของเซลล์ที่มีชีวิต รวมถึงเซลล์พืชที่ยังไม่ได้รับการศึกษาอย่างถี่ถ้วน มีไมโตคอนเดรียจำนวนมากในเซลล์ที่ต้องการพลังงานมากขึ้น
ไมโตคอนเดรียหายไป เซลล์พิเศษของอวัยวะสัตว์ประกอบด้วยไมโตคอนเดรียนับร้อยหรือนับพัน (สมอง หัวใจ กล้ามเนื้อ)
สารานุกรม YouTube
1 / 5
út นาโนเทอร์ไบน์ในเซลล์ สุดยอดเทคโนโลยีแห่งธรรมชาติ - การสังเคราะห์ ATP
√ เซลล์วิทยา การบรรยายครั้งที่ 17. ประจุไฟฟ้าและประจุในโมเลกุล
chain ห่วงโซ่การขนส่งอิเล็กตรอน
√ เซลล์วิทยา การบรรยายครั้งที่ 11. อันดูลิโพเดียม. อ็อกชไตน์ ไอ.แอล.
√ ทำไมคุณถึงอยากนอนและเกิดอะไรขึ้นกับร่างกายของคุณ ปิติพัฒน์.วี
คำบรรยาย
ต้นกำเนิดของไมโตคอนเดรีย
โครงสร้างไมโตคอนเดรีย
เยื่อหุ้มชั้นนอก
เยื่อหุ้มชั้นนอกของไมโตคอนเดรียมีความหนาประมาณ 7 นาโนเมตร ไม่ก่อให้เกิดการบุกรุกหรือรอยพับ และปิดอยู่บนตัวมันเอง เยื่อหุ้มชั้นนอกคิดเป็นประมาณ 7% ของพื้นที่ผิวของเยื่อหุ้มเซลล์ออร์แกเนลล์ทั้งหมด หน้าที่หลักคือแยกไมโตคอนเดรียออกจากไซโตพลาสซึม เยื่อหุ้มชั้นนอกของไมโตคอนเดรียประกอบด้วยไขมันที่สลับกับโปรตีน (อัตราส่วน 2:1) porin ซึ่งเป็นโปรตีนที่สร้างช่องมีบทบาทพิเศษโดยสร้างรูในเยื่อหุ้มชั้นนอกที่มีเส้นผ่านศูนย์กลาง 2-3 นาโนเมตรซึ่งโมเลกุลและไอออนขนาดเล็กที่มีน้ำหนักมากถึง 5 kDa สามารถทะลุผ่านได้ โมเลกุลขนาดใหญ่สามารถข้ามเยื่อหุ้มชั้นนอกผ่านการขนส่งแบบแอคทีฟผ่านโปรตีนขนส่งเมมเบรนแบบไมโตคอนเดรีย เมมเบรนด้านนอกมีลักษณะเป็นเอนไซม์: monooxygenase, acyl-CoA synthetase และ phospholipase A2 เยื่อหุ้มชั้นนอกของไมโตคอนเดรียสามารถโต้ตอบกับเยื่อหุ้มของเอนโดพลาสมิกเรติคูลัมได้ มีบทบาทสำคัญในการขนส่งไขมันและแคลเซียมไอออน
พื้นที่ระหว่างเมมเบรน
ช่องว่างระหว่างเมมเบรนคือช่องว่างระหว่างเยื่อหุ้มด้านนอกและด้านในของไมโตคอนเดรีย ความหนาของมันคือ 10-20 นาโนเมตร เนื่องจากเยื่อหุ้มชั้นนอกของไมโตคอนเดรียสามารถซึมผ่านไปยังโมเลกุลและไอออนขนาดเล็กได้ ความเข้มข้นของพวกมันในพื้นที่เพอริพลาสซึมจึงแตกต่างเล็กน้อยจากความเข้มข้นในไซโตพลาสซึม ในทางตรงกันข้าม โปรตีนขนาดใหญ่ต้องการเปปไทด์สัญญาณเฉพาะสำหรับการขนส่งจากไซโตพลาสซึมไปยังปริพลาสซึม ดังนั้นส่วนประกอบโปรตีนของปริพลาสซึมและไซโตพลาสซึมจึงแตกต่างกัน โปรตีนชนิดหนึ่งที่ไม่เพียงแต่มีอยู่ในเยื่อหุ้มชั้นในเท่านั้น แต่ยังอยู่ในช่องว่างรอบพลาสซึมด้วยคือไซโตโครม c
เยื่อหุ้มชั้นใน
ศักยภาพของพลังงาน (พลังงานสำรอง) ในโมเลกุลของยูบิควินอลนั้นต่ำกว่าในโมเลกุล NADH อย่างมีนัยสำคัญ และความแตกต่างของพลังงานดังกล่าวจะถูกเก็บไว้ชั่วคราวในรูปแบบของการไล่ระดับโปรตอนเคมีไฟฟ้า หลังเกิดขึ้นจากความจริงที่ว่าการถ่ายโอนอิเล็กตรอนผ่านกลุ่มเทียมของคอมเพล็กซ์ I ซึ่งนำไปสู่การลดศักยภาพพลังงานของอิเล็กตรอนนั้นมาพร้อมกับการถ่ายโอนเมมเบรนของโปรตอนสองตัวจากเมทริกซ์ไปยังช่องว่างระหว่างเมมเบรน ของไมโตคอนเดรีย
ยูบิควินอลที่ลดลงจะเคลื่อนที่ไปในระนาบของเมมเบรน ซึ่งไปถึงเอนไซม์ตัวที่สองของห่วงโซ่ระบบทางเดินหายใจ - คอมเพล็กซ์ III (ไซโตโครม ก่อนคริสต์ศักราช 1 - หลังเป็นไดเมอร์ที่มีน้ำหนักโมเลกุลมากกว่า 300 kDa เกิดจากโซ่โพลีเปปไทด์ 8 เส้นและมีอะตอมของเหล็กทั้งในรูปของศูนย์กลางของเหล็ก - ซัลเฟอร์และในรูปของเชิงซ้อนที่มีฮีม ข(ฉัน), ข(II) และ ค 1 - โมเลกุลเฮเทอโรไซคลิกเชิงซ้อนซึ่งมีอะตอมไนโตรเจน 4 อะตอมอยู่ที่มุมของสี่เหลี่ยมที่ยึดโลหะ Complex III เร่งปฏิกิริยาออกซิเดชันของยูบิควินอล 2 ชนิดไปเป็นยูบิควิโนน โดยลดโมเลกุลของไซโตโครม c 2 โมเลกุล (ตัวขนส่งที่มีฮีมอยู่ในช่องว่างระหว่างเมมเบรน) โปรตอนที่สี่ตัวที่ถูกแยกออกจากยูบิควินอลจะถูกปล่อยออกสู่ช่องว่างระหว่างเมมเบรน และจะก่อให้เกิดการไล่ระดับเคมีไฟฟ้าต่อไป
ขั้นตอนสุดท้ายถูกเร่งปฏิกิริยาโดย IV ที่ซับซ้อน (ไซโตโครม ค-ออกซิเดส) ที่มีน้ำหนักโมเลกุลประมาณ 200 kDa ประกอบด้วยสายโพลีเปปไทด์ 10-13 สาย และนอกเหนือจากฮีมที่แตกต่างกันสองชนิด ยังรวมถึงอะตอมของทองแดงหลายอะตอมที่จับกันอย่างแน่นหนากับโปรตีน ในกรณีนี้อิเล็กตรอนที่นำมาจากไซโตโครมรีดิวซ์ คเมื่อผ่านอะตอมของเหล็กและทองแดงในเชิงซ้อน IV พวกมันจะไปถึงออกซิเจนที่จับตัวอยู่ในศูนย์กลางที่แอคทีฟของเอนไซม์นี้ซึ่งนำไปสู่การก่อตัวของน้ำ
ดังนั้นปฏิกิริยาโดยรวมที่เร่งปฏิกิริยาโดยเอนไซม์ของลูกโซ่ทางเดินหายใจคือการออกซิเดชันของ NADH กับออกซิเจนเพื่อสร้างน้ำ โดยพื้นฐานแล้ว กระบวนการนี้ประกอบด้วยการถ่ายโอนอิเล็กตรอนแบบเป็นขั้นตอนระหว่างอะตอมของโลหะที่อยู่ในกลุ่มเทียมของโปรตีนเชิงซ้อนของห่วงโซ่ระบบทางเดินหายใจ โดยที่แต่ละคอมเพล็กซ์ที่ตามมาจะมีความสัมพันธ์ของอิเล็กตรอนสูงกว่ากลุ่มก่อนหน้า ในกรณีนี้ อิเล็กตรอนจะถูกถ่ายโอนไปตามสายโซ่จนกระทั่งรวมกับออกซิเจนโมเลกุลซึ่งมีความสัมพันธ์กับอิเล็กตรอนมากที่สุด พลังงานที่ปล่อยออกมาในกรณีนี้จะถูกเก็บไว้ในรูปแบบของการไล่ระดับเคมีไฟฟ้า (โปรตอน) ที่ทั้งสองด้านของเยื่อหุ้มไมโตคอนเดรียชั้นใน เชื่อกันว่าในระหว่างการขนส่งคู่อิเล็กตรอนผ่านห่วงโซ่ทางเดินหายใจจะมีการสูบโปรตอนสามถึงหกตัว
ขั้นตอนสุดท้ายของการทำงานของไมโตคอนเดรียคือการสร้าง ATP ซึ่งดำเนินการโดยโมเลกุลขนาดใหญ่พิเศษที่มีน้ำหนักโมเลกุล 500 kDa ที่สร้างไว้ในเยื่อหุ้มชั้นใน สารเชิงซ้อนนี้เรียกว่า ATP synthase กระตุ้นการสังเคราะห์ ATP โดยการแปลงพลังงานของการไล่ระดับเคมีไฟฟ้าของเมมเบรนของโปรตอนไฮโดรเจนให้เป็นพลังงานของพันธะพลังงานสูงของโมเลกุล ATP
เอทีพีซินเทส
ในแง่โครงสร้างและหน้าที่ ATP synthase ประกอบด้วยชิ้นส่วนขนาดใหญ่สองส่วน กำหนดด้วยสัญลักษณ์ F 1 และ F 0 ตัวแรก (ปัจจัยการมีเพศสัมพันธ์ F1) หันหน้าไปทางเมทริกซ์ไมโตคอนเดรียและยื่นออกมาจากเมมเบรนอย่างเห็นได้ชัดในรูปแบบของการก่อตัวทรงกลมสูง 8 นาโนเมตรและกว้าง 10 นาโนเมตร ประกอบด้วยหน่วยย่อยเก้าหน่วยที่แสดงโดยโปรตีนห้าประเภท สายโซ่โพลีเปปไทด์ของหน่วยย่อย α สามหน่วยและหน่วยย่อย β จำนวนเท่ากันนั้นจัดเรียงอยู่ในทรงกลมโปรตีนที่มีโครงสร้างคล้ายกัน ซึ่งรวมกันก่อตัวเป็นเฮกซาเมอร์ (αβ) 3 ซึ่งดูเหมือนลูกบอลที่แบนเล็กน้อย เช่นเดียวกับชิ้นส้มที่อัดแน่น หน่วยย่อย α และ β ที่ต่อเนื่องกันสร้างโครงสร้างที่โดดเด่นด้วยแกนสมมาตรลำดับที่สามที่มีมุมการหมุน 120° ที่ศูนย์กลางของเฮกซาเมอร์นี้คือหน่วยย่อย γ ซึ่งถูกสร้างขึ้นโดยสายโพลีเปปไทด์ที่ขยายออกไปสองเส้น และมีลักษณะคล้ายกับแท่งโค้งที่ผิดรูปเล็กน้อยยาวประมาณ 9 นาโนเมตร ในกรณีนี้ ส่วนล่างของหน่วยย่อย γ จะยื่นออกมาจากลูกบอลประมาณ 3 นาโนเมตร ไปทางเมมเบรนคอมเพล็กซ์ F0 นอกจากนี้ ที่อยู่ภายในเฮกซาเมอร์ก็คือหน่วยย่อย ε ย่อยที่เกี่ยวข้องกับ γ หน่วยย่อยสุดท้าย (ที่เก้า) ถูกกำหนดด้วยสัญลักษณ์ δ และตั้งอยู่ด้านนอกของ F 1
ส่วนเมมเบรนของ ATP synthase เรียกว่าคัปปลิ้งแฟกเตอร์ F0 นั้นเป็นโปรตีนเชิงซ้อนที่ไม่ชอบน้ำซึ่งแทรกซึมผ่านเมมเบรนและมีเฮมิแชนเนลสองช่องด้านในเพื่อให้ไฮโดรเจนโปรตอนผ่านไปได้ โดยรวมแล้ว F 0 complex ประกอบด้วยหน่วยย่อยโปรตีนหนึ่งหน่วย ก, สำเนาสองชุดของหน่วยย่อย ขรวมถึงสำเนาของหน่วยย่อยขนาดเล็ก 9 ถึง 12 ชุด ค- หน่วยย่อย ก(น้ำหนักโมเลกุล 20 kDa) ถูกแช่อยู่ในเมมเบรนโดยสมบูรณ์ โดยที่มันก่อตัวเป็นส่วน α-helical หกส่วนที่พาดผ่าน หน่วยย่อย ข(น้ำหนักโมเลกุล 30 kDa) มีเพียงบริเวณ α-helical ที่ค่อนข้างสั้นเพียงแห่งเดียวที่แช่อยู่ในเมมเบรน และส่วนที่เหลือจะยื่นออกมาจากเมมเบรนอย่างเห็นได้ชัดไปยัง F 1 และติดอยู่กับหน่วยย่อย δ ที่อยู่บนพื้นผิว หน่วยย่อยละ 9-12 ชุด ค(น้ำหนักโมเลกุล 6-11 kDa) เป็นโปรตีนที่ค่อนข้างเล็กของα-helices ที่ไม่ชอบน้ำสองตัวที่เชื่อมต่อถึงกันด้วยวง hydrophilic สั้น ๆ ที่มุ่งเน้นไปที่ F 1 และพวกมันรวมกันเป็นชุดเดียวในรูปของทรงกระบอกที่แช่อยู่ในเมมเบรน . หน่วยย่อย γ ที่ยื่นออกมาจากคอมเพล็กซ์ F 1 ไปทาง F 0 จะถูกจุ่มลงในกระบอกสูบนี้อย่างแม่นยำและติดแน่นกับมัน
ดังนั้นในโมเลกุล ATP synthase จึงสามารถแยกแยะหน่วยย่อยโปรตีนได้สองกลุ่ม ซึ่งสามารถเปรียบได้กับมอเตอร์สองส่วน: โรเตอร์และสเตเตอร์ “สเตเตอร์” นั้นไม่มีการเคลื่อนไหวสัมพันธ์กับเมมเบรน และรวมถึงเฮกซาเมอร์ทรงกลม (αβ) 3 ที่ตั้งบนพื้นผิวและหน่วยย่อย δ รวมถึงหน่วยย่อย กและ ขเมมเบรนคอมเพล็กซ์ F0. “โรเตอร์” ซึ่งเคลื่อนที่ได้สัมพันธ์กับโครงสร้างนี้ ประกอบด้วยหน่วยย่อย γ และ ε ซึ่งยื่นออกมาจากคอมเพล็กซ์ (αβ) 3 อย่างเห็นได้ชัด ซึ่งเชื่อมต่อกับวงแหวนของหน่วยย่อยที่แช่อยู่ในเมมเบรน ค.
ความสามารถในการสังเคราะห์ ATP เป็นคุณสมบัติของคอมเพล็กซ์เดี่ยว F 0 F 1 ซึ่งเกี่ยวข้องกับการถ่ายโอนโปรตอนไฮโดรเจนผ่าน F 0 ถึง F 1 ซึ่งในส่วนหลังจะมีศูนย์ตัวเร่งปฏิกิริยาที่แปลง ADP และฟอสเฟตเป็นโมเลกุล ATP . แรงผลักดันในการทำงานของ ATP synthase คือศักยภาพของโปรตอนที่สร้างขึ้นบนเยื่อหุ้มไมโตคอนเดรียชั้นในอันเป็นผลมาจากการทำงานของห่วงโซ่การขนส่งอิเล็กตรอน
แรงที่ขับเคลื่อน “โรเตอร์” ของ ATP synthase เกิดขึ้นเมื่อความต่างศักย์ระหว่างด้านนอกและด้านในของเมมเบรนสูงถึง > 220 mV และได้มาจากการไหลของโปรตอนที่ไหลผ่านช่องพิเศษใน F0 ซึ่งอยู่ที่ขอบเขตระหว่างหน่วยย่อย กและ ค- ในกรณีนี้ วิถีการถ่ายโอนโปรตอนประกอบด้วยองค์ประกอบทางโครงสร้างดังต่อไปนี้:
- "ครึ่งช่อง" ที่ไม่ได้อยู่โคแอกเซียลสองช่อง ช่องแรกช่วยให้แน่ใจว่ามีการจ่ายโปรตอนจากช่องว่างระหว่างเมมเบรนไปยังกลุ่มฟังก์ชันที่จำเป็น F0 และอีกช่องหนึ่งช่วยให้แน่ใจว่าพวกมันจะออกจากเมทริกซ์ไมโตคอนเดรีย
- วงแหวนของหน่วยย่อย คซึ่งแต่ละส่วนในส่วนกลางประกอบด้วยหมู่คาร์บอกซิลที่มีโปรตอน ซึ่งสามารถยึด H + จากช่องว่างระหว่างเมมเบรนและปล่อยพวกมันผ่านช่องโปรตอนที่เกี่ยวข้อง อันเป็นผลมาจากการกระจัดของหน่วยย่อยเป็นระยะ กับเกิดจากการไหลของโปรตอนผ่านช่องโปรตอน หน่วยย่อย γ หมุนไปจมอยู่ในวงแหวนของหน่วยย่อย กับ.
ดังนั้นกิจกรรมการเร่งปฏิกิริยาของ ATP synthase จึงเกี่ยวข้องโดยตรงกับการหมุนของ "โรเตอร์" ซึ่งการหมุนของหน่วยย่อย γ ทำให้เกิดการเปลี่ยนแปลงพร้อมกันในโครงสร้างของหน่วยย่อยตัวเร่งปฏิกิริยาทั้งสามหน่วย β ซึ่งท้ายที่สุดจะรับประกันการทำงานของเอนไซม์ . ในกรณีนี้ ในกรณีของการก่อตัวของ ATP "โรเตอร์" จะหมุนตามเข็มนาฬิกาด้วยความเร็วสี่รอบต่อวินาที และการหมุนนั้นเกิดขึ้นในการกระโดดแยกกันที่ 120° ซึ่งแต่ละอันจะมาพร้อมกับการก่อตัวของโมเลกุล ATP หนึ่งโมเลกุล .
ฟังก์ชันโดยตรงของการสังเคราะห์ ATP ได้รับการแปลในหน่วยย่อย β ของคอมเพล็กซ์คอนจูเกต F1 ในกรณีนี้ การกระทำแรกสุดในห่วงโซ่ของเหตุการณ์ที่นำไปสู่การก่อตัวของ ATP คือการจับกันของ ADP และฟอสเฟตกับศูนย์กลางที่ใช้งานของหน่วยย่อยβอิสระซึ่งอยู่ในสถานะ 1 เนื่องจากพลังงานของภายนอก แหล่งที่มา (กระแสโปรตอน) การเปลี่ยนแปลงโครงสร้างเกิดขึ้นใน F 1 คอมเพล็กซ์ซึ่งเป็นผลมาจากการที่ ADP และฟอสเฟตจับกันอย่างแน่นหนากับศูนย์ตัวเร่งปฏิกิริยา (สถานะ 2) ซึ่งการก่อตัวของพันธะโควาเลนต์ระหว่างพวกมันจะเป็นไปได้ การก่อตัวของเอทีพี ในขั้นตอนของ ATP synthase นี้ เอนไซม์แทบไม่ต้องใช้พลังงานเลย ซึ่งจะต้องใช้ในขั้นตอนต่อไปเพื่อปลดปล่อยโมเลกุล ATP ที่เกาะแน่นออกจากศูนย์กลางของเอนไซม์ ดังนั้นขั้นตอนต่อไปของการทำงานของเอนไซม์ก็คือ ผลจากการเปลี่ยนแปลงโครงสร้างที่ต้องอาศัยพลังงานในคอมเพล็กซ์ F 1 ทำให้หน่วยย่อย catalytic β-subunit ที่มีโมเลกุล ATP ที่ถูกผูกไว้อย่างแน่นหนาผ่านเข้าสู่สถานะ 3 ซึ่งการเชื่อมต่อของ ATP โดยมีศูนย์ตัวเร่งปฏิกิริยาอ่อนตัวลง ด้วยเหตุนี้โมเลกุล ATP จึงออกจากเอนไซม์และหน่วยย่อยβจะกลับสู่สถานะดั้งเดิม 1 ซึ่งช่วยให้มั่นใจในการหมุนเวียนของเอนไซม์
การทำงานของ ATP synthase เกี่ยวข้องกับการเคลื่อนไหวทางกลของแต่ละชิ้นส่วน ซึ่งทำให้สามารถจำแนกกระบวนการนี้เป็นปรากฏการณ์พิเศษที่เรียกว่า "ตัวเร่งปฏิกิริยาแบบหมุน" เช่นเดียวกับที่กระแสไฟฟ้าในขดลวดของมอเตอร์ไฟฟ้าขับเคลื่อนโรเตอร์โดยสัมพันธ์กับสเตเตอร์ การถ่ายโอนโปรตอนโดยตรงผ่าน ATP synthase ทำให้เกิดการหมุนของแต่ละหน่วยย่อยของปัจจัยการผัน F 1 สัมพันธ์กับหน่วยย่อยอื่น ๆ ของเอนไซม์ที่ซับซ้อน เช่น ผลจากการที่อุปกรณ์ผลิตพลังงานอันเป็นเอกลักษณ์นี้ทำงานทางเคมี - สังเคราะห์โมเลกุล ATP ต่อจากนั้น ATP จะเข้าสู่ไซโตพลาสซึมของเซลล์ ซึ่งถูกใช้ไปกับกระบวนการที่ขึ้นกับพลังงานที่หลากหลาย การถ่ายโอนดังกล่าวดำเนินการโดยเอนไซม์พิเศษ ATP/ADP translocase ที่สร้างขึ้นในเยื่อหุ้มไมโตคอนเดรีย ซึ่งแลกเปลี่ยน ATP ที่สังเคราะห์ขึ้นใหม่กับ ADP ของไซโตพลาสซึม ซึ่งรับประกันความปลอดภัยของแหล่งรวมอะดีนิลนิวคลีโอไทด์ภายในไมโตคอนเดรีย