- Mitochondria to maleńkie inkluzje w komórkach, które pierwotnie uważano za odziedziczone od bakterii. W większości komórek jest ich do kilku tysięcy, co stanowi od 15 do 50 procent objętości komórki. Są źródłem ponad 90 procent energii Twojego organizmu.
- Twoje mitochondria mają ogromny wpływ na zdrowie, zwłaszcza na raka, dlatego optymalizacja metabolizmu mitochondriów może być podstawą skutecznego leczenia raka
Rozmiar czcionki:
Od doktora Mercoli
Mitochondria: możesz nie wiedzieć, czym są, ale tak jest niezbędny dla Twojego zdrowia. Doktor Rhonda Patrick jest biomedykiem, która badała interakcje między metabolizmem mitochondriów, nieprawidłowym metabolizmem i rakiem.
Część jej pracy polega na identyfikacji wczesnych biomarkerów choroby. Na przykład uszkodzenie DNA jest wczesnym biomarkerem raka. Następnie próbuje określić, które mikroelementy pomagają naprawić uszkodzenia DNA.
Badała także funkcję i metabolizm mitochondriów, czym ostatnio się zainteresowałam. Jeśli po wysłuchaniu tego wywiadu chcesz dowiedzieć się więcej na ten temat, polecam zacząć od książki doktora Lee Knowa Life – The Epic Story of Our Mitochondria.
Mitochondria mają ogromny wpływ na zdrowie, zwłaszcza na raka, i zaczynam wierzyć, że optymalizacja metabolizmu mitochondriów może leżeć u podstaw skutecznego leczenia raka.
Znaczenie optymalizacji metabolizmu mitochondriów
Mitochondria to maleńkie organelle, które pierwotnie sądzono, że odziedziczyliśmy od bakterii. Prawie nie ma ich w czerwonych krwinkach i komórkach skóry, ale w komórkach rozrodczych jest ich 100 000, ale w większości komórek jest ich od jednego do 2000. Są one głównym źródłem energii dla twojego organizmu.
Aby narządy mogły prawidłowo funkcjonować, potrzebują energii, a energię tę wytwarzają mitochondria.
Ponieważ funkcja mitochondriów leży u podstaw wszystkiego, co dzieje się w organizmie, optymalizacja funkcji mitochondriów i zapobieganie dysfunkcjom mitochondriów poprzez pozyskiwanie wszystkich niezbędnych składników odżywczych i prekursorów wymaganych przez mitochondria jest niezwykle ważne dla zdrowia i zapobiegania chorobom.
Zatem jedną z uniwersalnych cech komórek nowotworowych jest poważne upośledzenie funkcji mitochondriów, w którym radykalnie zmniejsza się liczba funkcjonalnych mitochondriów.
Doktor Otto Warburg był lekarzem z dyplomem z chemii i bliskim przyjacielem Alberta Einsteina. Większość ekspertów uznaje Warburga za największego biochemika XX wieku.
W 1931 roku otrzymał Nagrodę Nobla za odkrycie, że komórki nowotworowe wykorzystują glukozę jako źródło produkcji energii. Nazywano to „efektem Warburga”, ale niestety zjawisko to nadal jest przez prawie wszystkich ignorowane.
Jestem przekonany, że dieta ketogenna, która radykalnie poprawia zdrowie mitochondriów, może pomóc w przypadku większości nowotworów, szczególnie w połączeniu z wychwytywaczem glukozy, jakim jest 3-bromopirogronian.
Jak mitochondria wytwarzają energię
Aby wytworzyć energię, mitochondria potrzebują tlenu z powietrza, którym oddychasz, oraz tłuszczu i glukozy z pożywienia, które spożywasz.
Te dwa procesy – oddychanie i jedzenie – są ze sobą powiązane w procesie zwanym fosforylacją oksydacyjną. Jest wykorzystywany przez mitochondria do wytwarzania energii w postaci ATP.
Mitochondria posiadają szereg łańcuchów transportu elektronów, poprzez które przenoszą elektrony ze zredukowanej formy pożywienia, które spożywasz, aby połączyć się z tlenem z powietrza, którym oddychasz, tworząc ostatecznie wodę.
Proces ten kieruje protony przez błonę mitochondrialną, ładując ATP (trifosforan adenozyny) z ADP (difosforan adenozyny). ATP transportuje energię po całym organizmie
Jednak w procesie tym powstają produkty uboczne, takie jak reaktywne formy tlenu (ROS), które szkoda komórek i mitochondrialnego DNA, następnie przenosząc je do DNA jądra komórkowego.
W ten sposób dochodzi do kompromisu. Wytwarzając energię, ciało starzeć się ze względu na destrukcyjne aspekty ROS, które powstają w tym procesie. Tempo starzenia się organizmu zależy w dużej mierze od tego, jak dobrze funkcjonują mitochondria i od ilości szkód, które można zrekompensować optymalizując dietę.
Rola mitochondriów w nowotworach
Kiedy pojawiają się komórki nowotworowe, reaktywne formy tlenu powstające jako produkt uboczny produkcji ATP wysyłają sygnał uruchamiający proces samobójstwa komórki, znany również jako apoptoza.
Ponieważ komórki nowotworowe powstają każdego dnia, jest to dobra rzecz. Zabijając uszkodzone komórki, organizm się ich pozbywa i zastępuje zdrowymi.
Komórki nowotworowe są jednak odporne na ten samobójczy protokół – mają przed nim wbudowaną obronę, jak wyjaśnili dr Warburg, a następnie Thomas Seyfried, który dogłębnie badał raka jako chorobę metaboliczną.
Jak wyjaśnia Patryk:
„Jednym z mechanizmów działania leków stosowanych w chemioterapii jest powstawanie reaktywnych form tlenu. Wyrządzają szkody, a to wystarczy, aby popchnąć komórkę nowotworową w stronę śmierci.
Myślę, że powodem tego jest to, że komórka nowotworowa, która nie wykorzystuje swoich mitochondriów, to znaczy nie wytwarza już reaktywnych form tlenu, i nagle zmusza się ją do korzystania z mitochondriów, w wyniku czego następuje przypływ reaktywnych form tlenu (w końcu tak robią mitochondria) i – bum, śmierć, bo komórka nowotworowa jest już gotowa na tę śmierć. Jest gotowa na śmierć.”
Dlaczego dobrze jest nie jeść wieczorem?
Od jakiegoś czasu jestem fanem postu przerywanego z różnych powodów, oczywiście ze względu na długowieczność i problemy zdrowotne, ale także dlatego, że wydaje się, że zapewnia on ogromne korzyści w zapobieganiu i leczeniu raka. Mechanizm tego jest związany z wpływem postu na mitochondria.
Jak wspomniano, głównym skutkiem ubocznym przenoszenia elektronów, w który zaangażowane są mitochondria, jest wyciek niektórych elektronów z łańcucha transportu elektronów i reakcja z tlenem, tworząc wolne rodniki ponadtlenkowe.
Anion ponadtlenkowy (powstający w wyniku redukcji tlenu o jeden elektron) jest prekursorem większości reaktywnych form tlenu i mediatorem utleniających reakcji łańcuchowych. Wolne rodniki tlenowe atakują lipidy w błonach komórkowych, receptorach białkowych, enzymach i DNA, co może przedwcześnie zabić mitochondria.
Niektóre wolne rodniki są wręcz pożyteczne, niezbędne organizmowi do regulowania funkcji komórkowych, jednak problemy pojawiają się przy nadmiernym tworzeniu się wolnych rodników. Niestety, z tego powodu u większości populacji zapada na większość chorób, w szczególności raka. Istnieją dwa sposoby rozwiązania tego problemu:
- Zwiększ ilość przeciwutleniaczy
- Zmniejsz produkcję wolnych rodników mitochondrialnych
Moim zdaniem jedną z najskuteczniejszych strategii redukcji wolnych rodników w mitochondriach jest ograniczenie ilości paliwa dostarczanego do organizmu. Nie jest to wcale kontrowersyjne, ponieważ ograniczenie kalorii konsekwentnie wykazuje wiele korzyści terapeutycznych. Jest to jeden z powodów, dla których post przerywany jest skuteczny, ponieważ ogranicza okres spożywania pokarmu, co automatycznie zmniejsza ilość spożywanych kalorii.
Jest to szczególnie skuteczne, jeśli nie jesz na kilka godzin przed snem, ponieważ jest to Twój najniższy stan metaboliczny.
To wszystko może wydawać się zbyt skomplikowane dla osób niebędących ekspertami, ale należy zrozumieć, że ponieważ organizm zużywa najmniej kalorii podczas snu, należy unikać jedzenia przed snem, ponieważ nadmiar paliwa w tym czasie doprowadzi do powstania nadmiernych ilości wolne rodniki, które niszczą tkanki, przyspieszają starzenie się i przyczyniają się do powstawania chorób przewlekłych.
Jak jeszcze post pomaga w zdrowym funkcjonowaniu mitochondriów?
Patrick zauważa również, że część mechanizmu odpowiedzialnego za skuteczność postu polega na tym, że organizm jest zmuszony pozyskiwać energię z lipidów i zapasów tłuszczu, co oznacza, że komórki są zmuszone korzystać z mitochondriów.
Mitochondria to jedyny mechanizm, dzięki któremu organizm może wytwarzać energię z tłuszczu. Zatem post pomaga aktywować mitochondria.
Uważa również, że odgrywa on ogromną rolę w mechanizmie, dzięki któremu przerywany post i dieta ketogenna zabijają komórki nowotworowe, i wyjaśnia, dlaczego niektóre leki aktywujące mitochondria mogą zabijać komórki nowotworowe. Ponownie dzieje się tak dlatego, że powstaje fala reaktywnych form tlenu, których uszkodzenie decyduje o wyniku sprawy, powodując śmierć komórek nowotworowych.
Odżywianie mitochondriów
Z żywieniowego punktu widzenia Patrick kładzie nacisk na następujące składniki odżywcze i ważne kofaktory niezbędne do prawidłowego funkcjonowania enzymów mitochondrialnych:
- Koenzym Q10 lub ubichinol (forma zredukowana)
- L-karnityna, która transportuje kwasy tłuszczowe do mitochondriów
- D-ryboza, która jest surowcem dla cząsteczek ATP
- Magnez
- Wszystkie witaminy z grupy B, w tym ryboflawina, tiamina i B6
- Kwas alfa liponowy (ALA)
Jak zauważa Patryk:
„Z różnych powodów wolę pozyskiwać jak najwięcej mikroelementów z pełnoziarnistej żywności. Po pierwsze, tworzą kompleks z błonnikami, co ułatwia ich wchłanianie.
Dodatkowo w tym przypadku zapewniony jest ich prawidłowy stosunek. Nie będziesz w stanie zdobyć ich w nadmiarze. Proporcje są dokładnie takie, jakich potrzebujesz. Istnieją inne elementy, które prawdopodobnie nie zostały jeszcze określone.
Musisz zachować szczególną czujność, upewniając się, że spożywasz szeroką gamę [pokarmów] i dostarczasz odpowiednich mikroelementów. Myślę, że przyjmowanie suplementu kompleksu witamin z grupy B jest pomocne z tego powodu.
Z tego powodu je akceptuję. Innym powodem jest to, że wraz z wiekiem nie wchłaniamy już tak łatwo witamin z grupy B, głównie ze względu na rosnącą sztywność błon komórkowych. Zmienia to sposób transportu witamin z grupy B do komórki. Są rozpuszczalne w wodzie, dzięki czemu nie odkładają się w tłuszczu. Nie da się nimi zatruć. W skrajnych przypadkach będziesz oddawać mocz nieco częściej. Ale jestem pewien, że są bardzo przydatne.”
Ćwiczenia mogą pomóc w utrzymaniu młodych mitochondriów
Ćwiczenia sprzyjają również zdrowiu mitochondriów, ponieważ pobudzają je do pracy. Jak wspomniano wcześniej, jednym ze skutków ubocznych zwiększonej aktywności mitochondriów jest powstawanie reaktywnych form tlenu, które pełnią rolę cząsteczek sygnalizacyjnych.
Jedną z sygnalizowanych przez nie funkcji jest tworzenie większej liczby mitochondriów. Kiedy więc ćwiczysz, organizm reaguje, tworząc więcej mitochondriów, aby zaspokoić zwiększone zapotrzebowanie na energię.
Starzenie się jest nieuniknione. Ale Twój wiek biologiczny może bardzo różnić się od wieku chronologicznego, a mitochondria mają wiele wspólnego ze starzeniem biologicznym. Patrick przytacza najnowsze badania, które pokazują, w jaki sposób ludzie mogą się starzeć biologicznie Bardzo w różnym tempie.
Naukowcy zmierzyli kilkanaście różnych biomarkerów, takich jak długość telomerów, uszkodzenie DNA, cholesterol LDL, metabolizm glukozy i wrażliwość na insulinę, w trzech momentach życia ludzi: w wieku 22, 32 i 38 lat.
„Odkryliśmy, że na podstawie markerów biologicznych osoba w wieku 38 lat może biologicznie wyglądać o 10 lat młodziej lub starzej. Mimo tego samego wieku, starzenie się biologiczne następuje w zupełnie innym tempie.
Co ciekawe, kiedy fotografowano te osoby, a ich zdjęcia pokazywano przechodniom i proszono o odgadnięcie wieku chronologicznego przedstawionych osób, ludzie odgadywali wiek biologiczny, a nie chronologiczny”.
Zatem niezależnie od tego, ile masz lat, wygląd odpowiada Twoim biologicznym biomarkerom, które w dużej mierze zależą od stanu mitochondriów. Chociaż starzenia się nie można uniknąć, masz dużą kontrolę nad tym, jak się starzejesz, a to daje dużo mocy. Jednym z kluczowych czynników jest utrzymanie mitochondriów w dobrym stanie.
Według Patricka „młodość” to nie tyle wiek chronologiczny, ile to, ile lat się czujesz i jak dobrze funkcjonuje twoje ciało:
„Chcę wiedzieć, jak zoptymalizować swoją wydajność umysłową i wyniki sportowe. Chcę przedłużyć swoją młodość. Chcę dożyć 90 lat. A kiedy to zrobię, chcę surfować w San Diego w taki sam sposób, w jaki robiłem to, gdy miałem 20 lat. Szkoda, że nie zniknąłem tak szybko, jak niektórzy ludzie. Lubię opóźniać ten upadek i jak najdłużej przedłużać młodość, abym mógł jak najdłużej cieszyć się życiem.”
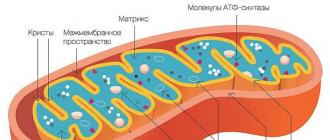
Struktura. Aparat powierzchniowy mitochondriów składa się z dwóch błon - zewnętrznej i wewnętrznej. Zewnętrzna męmbrana gładka, oddziela mitochondria od hialoplazmy. Poniżej znajduje się zgięcie wewnętrzna membrana, które tworzy Christie(grzbiety). Po obu stronach cristae znajdują się małe ciała w kształcie grzybów zwane oksysomami lub ATP-somami. Zawierają enzymy biorące udział w fosforylacji oksydacyjnej (dodawanie reszt fosforanowych do ADP w celu utworzenia ATP). Liczba cristae w mitochondriach jest związana z potrzebami energetycznymi komórki; w szczególności w komórkach mięśniowych mitochondria zawierają bardzo dużą liczbę cristae. Wraz ze zwiększoną funkcją komórek mitochondria stają się bardziej owalne lub wydłużone, a liczba cristae wzrasta.
Mitochondria mają swój własny genom, ich rybosomy typu 70S różnią się od rybosomów cytoplazmy. DNA mitochondrialny ma głównie postać cykliczną (plazmidy), koduje wszystkie trzy typy własnego RNA i dostarcza informacji do syntezy niektórych białek mitochondrialnych (około 9%). Zatem mitochondria można uznać za organelle półautonomiczne. Mitochondria to samoreplikujące się (zdolne do reprodukcji) organelle. Odnowa mitochondriów zachodzi podczas całego cyklu komórkowego. Przykładowo w komórkach wątroby są one zastępowane nowymi po niemal 10 dniach. Za najbardziej prawdopodobny sposób reprodukcji mitochondriów uważa się ich podział: w środku mitochondriów pojawia się zwężenie lub pojawia się przegroda, po czym organelle dzielą się na dwa nowe mitochondria. Mitochondria powstają z promitochondriów - okrągłych ciał o średnicy do 50 nm z podwójną błoną.
Funkcje . Mitochondria biorą udział w procesach energetycznych komórki, zawierają enzymy związane z wytwarzaniem energii i oddychaniem komórkowym. Innymi słowy, mitochondrium jest rodzajem biochemicznej mini-fabryki, która przekształca energię związków organicznych w zastosowaną energię ATP. W mitochondriach proces energetyczny rozpoczyna się w matrixie, gdzie następuje rozkład kwasu pirogronowego w cyklu Krebsa. Podczas tego procesu atomy wodoru są uwalniane i transportowane przez łańcuch oddechowy. Uwolniona w tym przypadku energia jest wykorzystywana w kilku częściach łańcucha oddechowego do przeprowadzenia reakcji fosforylacji - syntezy ATP, czyli dodania grupy fosforanowej do ADP. Dzieje się tak na wewnętrznej błonie mitochondriów. Więc, funkcja energetyczna mitochondria integrują się z: a) utlenianiem związków organicznych zachodzącym w matrixie, dzięki czemu nazywane są mitochondria ośrodek oddechowy komórek b) Synteza ATP odbywa się na cristae, dzięki czemu nazywane są mitochondria stacje energetyczne komórek. Ponadto mitochondria biorą udział w regulacji metabolizmu wody, odkładaniu jonów wapnia, produkcji prekursorów hormonów steroidowych, metabolizmie (np. mitochondria w komórkach wątroby zawierają enzymy, które pozwalają im neutralizować amoniak) i innych.
BIOLOGIA + Choroby mitochondrialne to grupa chorób dziedzicznych związanych z defektami mitochondriów, które prowadzą do upośledzenia oddychania komórkowego. Są przenoszone przez linię żeńską na dzieci obu płci, ponieważ jajo ma większą objętość cytoplazmy i odpowiednio przekazuje potomkom większą liczbę mitochondriów. DNA mitochondrialny, w przeciwieństwie do DNA jądrowego, nie jest chroniony przez białka histonowe, a mechanizmy naprawcze odziedziczone od bakterii przodków są niedoskonałe. Dlatego mutacje kumulują się w mitochondrialnym DNA 10-20 razy szybciej niż w DNA jądrowym, co prowadzi do chorób mitochondrialnych. We współczesnej medycynie znanych jest obecnie około 50 z nich, na przykład zespół chronicznego zmęczenia, migrena, zespół Bartha, zespół Pearsona i wiele innych.
Mitochondria to mikroskopijne organelle związane z błoną, które dostarczają komórce energię. Dlatego nazywane są stacjami energetycznymi (bateriami) ogniw.
Mitochondria nie występują w komórkach organizmów prostych, bakterii i entameb, które żyją bez użycia tlenu. Niektóre zielone algi, trypanosomy, zawierają jedno duże mitochondrium, a komórki mięśnia sercowego i mózgu mają od 100 do 1000 takich organelli.
Cechy konstrukcyjne
Mitochondria to organelle dwubłonowe; mają błonę zewnętrzną i wewnętrzną, przestrzeń międzybłonową między nimi oraz macierz.
Zewnętrzna męmbrana. Jest gładka, nie ma fałd i oddziela zawartość wewnętrzną od cytoplazmy. Jego szerokość wynosi 7 nm i zawiera lipidy i białka. Ważną rolę odgrywa porina, białko tworzące kanały w błonie zewnętrznej. Zapewniają wymianę jonową i molekularną.
Przestrzeń międzybłonowa. Rozmiar przestrzeni międzybłonowej wynosi około 20 nm. Wypełniająca ją substancja ma podobny skład do cytoplazmy, z wyjątkiem dużych cząsteczek, które mogą tu przedostać się jedynie poprzez transport aktywny.

Wewnętrzna membrana. Zbudowany jest głównie z białka, tylko jedna trzecia przypada na substancje lipidowe. Duża liczba białek to białka transportowe, ponieważ w błonie wewnętrznej brakuje swobodnie przepuszczalnych porów. Tworzy wiele odrostów - cristae, które wyglądają jak spłaszczone grzbiety. Utlenianie związków organicznych do CO 2 w mitochondriach zachodzi na błonach cristae. Proces ten jest zależny od tlenu i zachodzi pod działaniem syntetazy ATP. Uwolniona energia jest magazynowana w postaci cząsteczek ATP i wykorzystywana w miarę potrzeb.
Matryca– środowisko wewnętrzne mitochondriów ma ziarnistą, jednorodną strukturę. W mikroskopie elektronowym można zobaczyć granulki i włókna w kulkach swobodnie leżących pomiędzy cristae. W matrixie znajduje się półautonomiczny układ syntezy białek - znajdują się tu DNA, wszystkie rodzaje RNA i rybosomy. Mimo to większość białek pochodzi z jądra, dlatego mitochondria nazywane są organellami półautonomicznymi.
Lokalizacja i podział komórki
Hondriom to grupa mitochondriów skupiona w jednej komórce. Są one różnie zlokalizowane w cytoplazmie, co zależy od specjalizacji komórek. Umieszczenie w cytoplazmie zależy również od otaczających organelli i inkluzji. W komórkach roślinnych zajmują obrzeża, ponieważ mitochondria są wypychane w kierunku błony przez centralną wakuolę. W komórkach nabłonka nerek błona tworzy wypustki, pomiędzy którymi znajdują się mitochondria.
W komórkach macierzystych, gdzie energia jest zużywana równomiernie przez wszystkie organelle, mitochondria są rozmieszczone chaotycznie. W wyspecjalizowanych ogniwach skupiają się one głównie w obszarach o największym zużyciu energii. Na przykład w mięśniach prążkowanych znajdują się one w pobliżu miofibryli. W plemnikach pokrywają one spiralnie oś wici, ponieważ do jej wprawienia i poruszenia plemnikiem potrzeba dużo energii. Pierwotniaki poruszające się za pomocą rzęsek również zawierają u podstawy dużą liczbę mitochondriów.
Dział. Mitochondria są zdolne do niezależnej reprodukcji, posiadając własny genom. Organelle są podzielone przewężeniami lub przegrodami. Częstotliwość powstawania nowych mitochondriów w różnych komórkach jest różna, np. w tkance wątroby są one wymieniane co 10 dni.
Funkcje w komórce
- Główną funkcją mitochondriów jest tworzenie cząsteczek ATP.
- Odkładanie jonów wapnia.
- Udział w wymianie wody.
- Synteza prekursorów hormonów steroidowych.
Biologia molekularna to nauka badająca rolę mitochondriów w metabolizmie. Przekształcają także pirogronian w acetylokoenzym A i beta-oksydację kwasów tłuszczowych.
| Tabela: budowa i funkcje mitochondriów (w skrócie) | ||
|---|---|---|
| Elementy konstrukcyjne | Struktura | Funkcje |
| Zewnętrzna męmbrana | Gładka skorupa zbudowana z lipidów i białek | Oddziela zawartość wewnętrzną od cytoplazmy |
| Przestrzeń międzybłonowa | Są jony wodoru, białka, mikrocząsteczki | Tworzy gradient protonowy |
| Wewnętrzna membrana | Tworzy wypustki - cristae, zawiera systemy transportu białek | Transfer makrocząsteczek, utrzymanie gradientu protonów |
| Matryca | Lokalizacja enzymów cyklu Krebsa, DNA, RNA, rybosomów | Utlenianie tlenowe z uwolnieniem energii, konwersja pirogronianu do acetylokoenzymu A. |
| Rybosomy | Połączone dwie podjednostki | Synteza białek |
Podobieństwa między mitochondriami i chloroplastami
Wspólne właściwości mitochondriów i chloroplastów wynikają przede wszystkim z obecności podwójnej błony.
Oznaki podobieństwa obejmują także zdolność do samodzielnej syntezy białka. Organelle te mają własne DNA, RNA i rybosomy.
Zarówno mitochondria, jak i chloroplasty mogą dzielić się przez zwężenie.
Łączy je również zdolność do wytwarzania energii; mitochondria są bardziej wyspecjalizowane w tej funkcji, ale chloroplasty wytwarzają również cząsteczki ATP podczas procesów fotosyntezy. Zatem komórki roślinne mają mniej mitochondriów niż komórki zwierzęce, ponieważ chloroplasty częściowo pełnią za nie funkcje.
Opiszmy pokrótce podobieństwa i różnice:
- Są to organelle dwubłonowe;
- wewnętrzna błona tworzy wypukłości: cristae są charakterystyczne dla mitochondriów, a thillacoidy są charakterystyczne dla chloroplastów;
- mają swój własny genom;
- zdolne do syntezy białek i energii.
Organelle te różnią się funkcjami: mitochondria służą do syntezy energii, zachodzi tu oddychanie komórkowe, chloroplasty są potrzebne komórkom roślinnym do fotosyntezy.
Mitochondria (z greckiego μίτος (mitos) - nić i χονδρίον (chondrion) - granulka) to komórkowa organella z dwiema błonami, która zawiera własny materiał genetyczny, mitochondrialny. Występują jako kuliste lub rurkowate struktury komórkowe u prawie wszystkich eukariontów, ale nie u prokariotów.
Mitochondria to organelle, które regenerują wysokoenergetyczną cząsteczkę adenozynotrifosforanu w łańcuchu oddechowym. Oprócz tej fosforylacji oksydacyjnej pełnią inne ważne zadania, m.in. biorą udział w tworzeniu klastrów żelaza i siarki. Strukturę i funkcje takich organelli omówiono szczegółowo poniżej.
W kontakcie z
Informacje ogólne
Szczególnie dużo mitochondriów znajduje się na obszarach o wysokim zużyciu energii. Należą do nich mięśnie, nerwy, komórki czuciowe i oocyty. W strukturach komórkowych mięśnia sercowego udział objętościowy tych organelli sięga 36%. Mają średnicę około 0,5-1,5 mikrona i różnorodne kształty, od kul po skomplikowane gwinty. Ich ilość dobierana jest z uwzględnieniem potrzeb energetycznych ogniwa.
Komórki eukariotyczne, które tracą mitochondria nie można ich przywrócić. Istnieją również eukarionty bez nich, na przykład niektóre pierwotniaki. Liczba tych organelli na jednostkę komórkową wynosi zwykle od 1000 do 2000, a udział objętościowy wynosi 25%. Ale wartości te mogą się znacznie różnić w zależności od rodzaju struktury komórkowej i organizmu. W dojrzałym plemniku jest ich około czterech do pięciu, a w dojrzałym jajku kilkaset tysięcy.
Mitochondria przenoszone są przez plazmę komórki jajowej wyłącznie od matki, co było powodem badania linii matczynych. Obecnie ustalono, że również poprzez plemniki niektóre organelle męskie są importowane do osocza zapłodnionego jaja (zygoty). Prawdopodobnie zostaną one rozwiązane dość szybko. Istnieje jednak kilka przypadków, w których lekarzom udało się udowodnić, że mitochondria dziecka pochodzą z linii ojcowskiej. Choroby spowodowane mutacjami w genach mitochondrialnych dziedziczone są wyłącznie od matki.
Ciekawy! Popularny naukowy termin „potęga komórki” został ukuty w 1957 roku przez Philipa Sikiewitz.
Schemat struktury mitochondriów
Rozważmy cechy strukturalne tych ważnych struktur. Powstają w wyniku połączenia kilku elementów. Powłoka tych organelli składa się z błony zewnętrznej i wewnętrznej, one z kolei składają się z dwuwarstw fosfolipidowych i białek. Obie muszle różnią się swoimi właściwościami. Pomiędzy nimi znajduje się pięć różnych przedziałów: błona zewnętrzna, przestrzeń międzybłonowa (przestrzeń między dwiema błonami), błona wewnętrzna, crista i matrix (przestrzeń wewnątrz błony wewnętrznej), ogólnie - wewnętrzne struktury organelli .
Na ilustracjach w podręcznikach mitochondrium wygląda przeważnie jak oddzielna organella w kształcie fasoli. Czy to naprawdę? Nie, tworzą się kanalikowa sieć mitochondriów, który może przejść i zmienić całą jednostkę komórkową. Mitochondria w komórce są zdolne do łączenia (poprzez fuzję) i ponownego podziału (poprzez rozszczepienie).
Notatka! U drożdży w ciągu jednej minuty zachodzą około dwie fuzje mitochondriów. Dlatego niemożliwe jest dokładne określenie aktualnej liczby mitochondriów w komórkach.
Zewnętrzna męmbrana
 Zewnętrzna powłoka otacza całą organellę i zawiera kanały kompleksów białkowych, które umożliwiają wymianę cząsteczek i jonów pomiędzy mitochondrium a cytozolem. Duże cząsteczki nie może przejść przez membranę.
Zewnętrzna powłoka otacza całą organellę i zawiera kanały kompleksów białkowych, które umożliwiają wymianę cząsteczek i jonów pomiędzy mitochondrium a cytozolem. Duże cząsteczki nie może przejść przez membranę.
Zewnętrzna, która obejmuje całą organellę i nie jest pofałdowana, ma stosunek wagowy fosfolipidów do białek wynoszący 1:1, a zatem jest podobna do błony komórkowej eukariotów. Zawiera wiele integralnych białek, porin. Poryny tworzą kanały umożliwiające swobodną dyfuzję cząsteczek o masie do 5000 daltonów przez membranę. Większe białka mogą dokonać inwazji, gdy sekwencja sygnałowa na końcu N wiąże się z dużą podjednostką białka transloksazy, z której następnie aktywnie przemieszczają się wzdłuż otoczki błony.
Jeśli w błonie zewnętrznej wystąpią pęknięcia, białka z przestrzeni międzybłonowej mogą przedostać się do cytozolu, który może prowadzić do śmierci komórki. Błona zewnętrzna może połączyć się z błoną siateczki śródplazmatycznej i następnie utworzyć strukturę zwaną MAM (ER związany z mitochondriami). Jest ważny dla sygnalizacji między ER a mitochondrium, co jest również niezbędne do transportu.
Przestrzeń międzybłonowa
Obszar ten to szczelina pomiędzy błoną zewnętrzną i wewnętrzną. Ponieważ zewnętrzna umożliwia swobodną penetrację małych cząsteczek, ich stężenie, np. jonów i cukrów, w przestrzeni międzybłonowej jest identyczne ze stężeniami w cytozolu. Jednakże duże białka wymagają przekazania określonej sekwencji sygnałowej, przez co skład białek różni się pomiędzy przestrzenią międzybłonową a cytozolem. Zatem białkiem zatrzymywanym w przestrzeni międzybłonowej jest cytochrom.
Wewnętrzna membrana
Wewnętrzna błona mitochondrialna zawiera białka spełniające cztery rodzaje funkcji:
- Białka – realizują reakcje utleniania łańcucha oddechowego.
- Syntaza adenozynotrójfosforanu, która wytwarza ATP w matrixie.
- Specyficzne białka transportowe, które regulują przejście metabolitów między macierzą a cytoplazmą.
- Systemy importu białek.
Wewnętrzny zawiera w szczególności podwójny fosfolipid, kardiolipinę, zastąpiony czterema kwasami tłuszczowymi. Kardiolipina powszechnie występuje w błonach mitochondriów i błonach plazmatycznych bakterii. Występuje głównie w organizmie człowieka w obszarach o dużej aktywności metabolicznej lub aktywność o wysokiej energii, taka jak kurczliwe kardiomiocyty w mięśniu sercowym.
Uwaga! Błona wewnętrzna zawiera ponad 150 różnych polipeptydów, co stanowi około 1/8 wszystkich białek mitochondrialnych. W rezultacie stężenie lipidów jest niższe niż w zewnętrznej dwuwarstwie, a jej przepuszczalność jest mniejsza.
 Podzielone na liczne cristae, rozszerzają zewnętrzny obszar wewnętrznej błony mitochondrialnej, zwiększając jej zdolność do wytwarzania ATP.
Podzielone na liczne cristae, rozszerzają zewnętrzny obszar wewnętrznej błony mitochondrialnej, zwiększając jej zdolność do wytwarzania ATP.
Na przykład w typowych mitochondriach wątroby obszar zewnętrzny, szczególnie cristae, jest około pięciokrotnie większy od obszaru błony zewnętrznej. Stacje energetyczne komórek mających większe zapotrzebowanie na ATP, m.in. komórki mięśniowe zawierają więcej cristae, niż typowe mitochondria wątroby.
Wewnętrzna powłoka otacza matrix, wewnętrzny płyn mitochondriów. Odpowiada cytozolowi bakterii i zawiera mitochondrialny DNA, enzymy cyklu cytrynianowego i ich własne mitochondrialne rybosomy, które różnią się od rybosomów w cytozolu (ale także od bakterii). Przestrzeń międzybłonowa zawiera enzymy, które mogą fosforylować nukleotydy poprzez zużycie ATP.
Funkcje
- Ważne ścieżki degradacji: cykl cytrynianowy, podczas którego pirogronian jest wprowadzany z cytozolu do matrix. Pirogronian jest następnie dekarboksylowany przez dehydrogenazę pirogronianową do acetylokoenzymu A. Innym źródłem acetylokoenzymu A jest degradacja kwasów tłuszczowych (β-oksydacja), która zachodzi w komórkach zwierzęcych w mitochondriach, natomiast w komórkach roślinnych jedynie w glioksysomach i peroksysomach. W tym celu acylo-koenzym A jest przenoszony z cytozolu poprzez wiązanie się z karnityną przez wewnętrzną błonę mitochondrialną i przekształcany w acetylokoenzym A. Stąd większość redukujących równoważników w cyklu Krebsa (znanym również jako cykl Krebsa lub cykl kwasu trikarboksylowego), które następnie są przekształcane w ATP w łańcuchu oksydacyjnym.
- Łańcuch oksydacyjny. Pomiędzy przestrzenią międzybłonową a macierzą mitochondrialną ustalono gradient elektrochemiczny, który służy do produkcji ATP przy użyciu syntazy ATP, poprzez procesy przenoszenia elektronów i akumulacji protonów. Otrzymuje się elektrony i protony potrzebne do wytworzenia gradientu poprzez degradację oksydacyjną ze składników odżywczych(takie jak glukoza) wchłaniane przez organizm. Glikoliza początkowo zachodzi w cytoplazmie.
- Apoptoza (programowana śmierć komórki)
- Magazynowanie wapnia: poprzez zdolność wchłaniania jonów wapnia, a następnie ich uwalniania, mitochondria zakłócają homeostazę komórek.
- Synteza klastrów żelazowo-siarkowych wymagana jest m.in. przez wiele enzymów łańcucha oddechowego. Funkcja ta jest obecnie uważana za podstawową funkcję mitochondriów, tj. ponieważ jest to powód, dla którego prawie wszystkie komórki, aby przetrwać, polegają na stacjach energetycznych.
![]()
Matryca
Jest to przestrzeń zawarta w wewnętrznej błonie mitochondrialnej. Zawiera około dwóch trzecich całkowitego białka. Odgrywa kluczową rolę w produkcji ATP poprzez syntazę ATP, zawartą w błonie wewnętrznej. Zawiera wysoce skoncentrowaną mieszaninę setek różnych enzymów (głównie biorących udział w degradacji kwasów tłuszczowych i pirogronianu), rybosomy specyficzne dla mitochondriów, informacyjny RNA i kilka kopii DNA genomu mitochondrialnego.
Organelle te mają swój własny genom, a także niezbędny do tego sprzęt enzymatyczny przeprowadzając własną biosyntezę białek.
Mitochondria Co to jest mitochondria i jakie są ich funkcje
Budowa i funkcjonowanie mitochondriów
Wniosek
Zatem mitochondria nazywane są elektrowniami komórkowymi, które wytwarzają energię i zajmują wiodące miejsce w życiu i przetrwaniu w szczególności pojedynczej komórki i ogólnie żywego organizmu. Mitochondria są integralną częścią żywej komórki, w tym komórek roślinnych, które nie zostały jeszcze w pełni zbadane. Szczególnie dużo mitochondriów znajduje się w tych komórkach, które wymagają więcej energii.
Mitochondria są nieobecne. Wyspecjalizowane komórki narządów zwierzęcych zawierają setki, a nawet tysiące mitochondriów (mózg, serce, mięśnie).
Encyklopedyczny YouTube
1 / 5
✪ Nanoturbiny w komórkach. SUPER TECHNOLOGIA NATURY - synteza ATP.
✪ Cytologia. Wykład 17. Ładunki elektryczne i ładunki w cząsteczkach
✪ Łańcuch transportu elektronów
✪ Cytologia. Wykład 11. Undulipodium. Okshtein I.L.
✪ Dlaczego chcesz spać i co dzieje się z Twoim ciałem. Pyatibrat.V
Napisy na filmie obcojęzycznym
Pochodzenie mitochondriów
Struktura mitochondriów
Zewnętrzna męmbrana
Zewnętrzna błona mitochondriów ma grubość około 7 nm, nie tworzy wgłębień ani fałdów i jest zamknięta. Błona zewnętrzna stanowi około 7% powierzchni wszystkich błon organelli komórkowych. Główną funkcją jest oddzielenie mitochondriów od cytoplazmy. Zewnętrzna błona mitochondrium składa się z lipidów przeplatanych białkami (stosunek 2:1). Szczególną rolę odgrywa porina, białko kanałotwórcze: tworzy w błonie zewnętrznej dziury o średnicy 2-3 nm, przez które mogą przenikać małe cząsteczki i jony o masie do 5 kDa. Duże cząsteczki mogą przenikać przez błonę zewnętrzną jedynie poprzez aktywny transport za pośrednictwem białek transportujących błonę mitochondrialną. Błonę zewnętrzną charakteryzuje obecność enzymów: monooksygenazy, syntetazy acylo-CoA i fosfolipazy A2. Zewnętrzna błona mitochondrium może oddziaływać z błoną retikulum endoplazmatycznego; odgrywa ważną rolę w transporcie lipidów i jonów wapnia.
Przestrzeń międzybłonowa
Przestrzeń międzybłonowa to przestrzeń pomiędzy zewnętrzną i wewnętrzną błoną mitochondrium. Jego grubość wynosi 10-20 nm. Ponieważ zewnętrzna błona mitochondrium jest przepuszczalna dla małych cząsteczek i jonów, ich stężenie w przestrzeni peryplazmatycznej niewiele różni się od tego w cytoplazmie. Wręcz przeciwnie, duże białka wymagają specyficznych peptydów sygnałowych do transportu z cytoplazmy do przestrzeni peryplazmatycznej; dlatego składniki białkowe przestrzeni peryplazmatycznej i cytoplazmy są różne. Jednym z białek znajdujących się nie tylko w błonie wewnętrznej, ale także w przestrzeni peryplazmatycznej jest cytochrom.
Wewnętrzna membrana
Potencjał energetyczny (rezerwa energii) w cząsteczce ubichinolu jest znacznie niższy niż w cząsteczce NADH, a różnica tej energii jest chwilowo magazynowana w postaci elektrochemicznego gradientu protonów. To ostatnie powstaje w wyniku tego, że przenoszeniu elektronów przez grupy prostetyczne kompleksu I, prowadzącemu do zmniejszenia potencjału energetycznego elektronów, towarzyszy transbłonowe przeniesienie dwóch protonów z matrycy do przestrzeni międzybłonowej mitochondrium.
Zredukowany ubichinol migruje w płaszczyźnie błony, gdzie dociera do drugiego enzymu łańcucha oddechowego – kompleksu III (cytochromu pne 1 ). Ten ostatni jest dimerem o masie cząsteczkowej powyżej 300 kDa, zbudowanym z ośmiu łańcuchów polipeptydowych i zawierającym atomy żelaza zarówno w postaci centrów żelazowo-siarkowych, jak i w postaci kompleksów z hemami B(I), B(II) i C 1 - złożone cząsteczki heterocykliczne z czterema atomami azotu umieszczonymi w rogach kwadratu wiążącego metal. Kompleks III katalizuje utlenianie dwóch ubichinoli do ubichinonów, redukując dwie cząsteczki cytochromu c (transporter zawierający hem zlokalizowany w przestrzeni międzybłonowej). Cztery protony oddzielone od ubichinoli są uwalniane do przestrzeni międzybłonowej, kontynuując tworzenie gradientu elektrochemicznego.
Ostatni etap jest katalizowany przez kompleks IV (cytochrom C-oksydaza) o masie cząsteczkowej około 200 kDa, składającej się z 10-13 łańcuchów polipeptydowych i oprócz dwóch różnych hemów, zawierającej także kilka atomów miedzi ściśle związanych z białkami. W tym przypadku elektrony są pobierane ze zredukowanego cytochromu C przechodząc przez atomy żelaza i miedzi w kompleksie IV, docierają do tlenu związanego w centrum aktywnym tego enzymu, co prowadzi do powstania wody.
Zatem ogólną reakcją katalizowaną przez enzymy łańcucha oddechowego jest utlenianie NADH tlenem z wytworzeniem wody. Zasadniczo proces ten polega na stopniowym przenoszeniu elektronów pomiędzy atomami metali obecnymi w grupach prostetycznych kompleksów białkowych łańcucha oddechowego, gdzie każdy kolejny kompleks ma większe powinowactwo elektronowe niż poprzedni. W tym przypadku same elektrony są przenoszone wzdłuż łańcucha, aż łączą się z tlenem cząsteczkowym, który ma największe powinowactwo do elektronów. Uwolniona w tym przypadku energia jest magazynowana w postaci gradientu elektrochemicznego (protonowego) po obu stronach wewnętrznej błony mitochondrialnej. Uważa się, że podczas transportu par elektronów przez łańcuch oddechowy pompowanych jest od trzech do sześciu protonów.
Ostatnim etapem funkcjonowania mitochondriów jest wytwarzanie ATP, realizowane przez specjalny kompleks makromolekularny o masie cząsteczkowej 500 kDa wbudowany w błonę wewnętrzną. Kompleks ten, zwany syntazą ATP, katalizuje syntezę ATP poprzez konwersję energii transbłonowego gradientu elektrochemicznego protonów wodoru w energię wysokoenergetycznego wiązania cząsteczki ATP.
Syntaza ATP
Pod względem strukturalnym i funkcjonalnym syntaza ATP składa się z dwóch dużych fragmentów, oznaczonych symbolami F 1 i F 0. Pierwszy z nich (czynnik sprzęgający F1) jest zwrócony w stronę matrix mitochondrialnej i wyraźnie wystaje z błony w postaci kulistej formacji o wysokości 8 nm i szerokości 10 nm. Składa się z dziewięciu podjednostek reprezentowanych przez pięć rodzajów białek. Łańcuchy polipeptydowe trzech podjednostek α i takiej samej liczby podjednostek β ułożone są w globulki białkowe o podobnej budowie, które razem tworzą heksamer (αβ) 3, który wygląda jak lekko spłaszczona kula. Podobnie jak ciasno upakowane plasterki pomarańczy, kolejne podjednostki α i β tworzą strukturę charakteryzującą się osią symetrii trzeciego rzędu z kątem obrotu 120°. W centrum tego heksameru znajduje się podjednostka γ, która jest utworzona przez dwa wydłużone łańcuchy polipeptydowe i przypomina lekko zdeformowany zakrzywiony pręt o długości około 9 nm. W tym przypadku dolna część podjednostki γ wystaje z kuli na 3 nm w stronę kompleksu błonowego F0. W heksamerze znajduje się również mniejsza podjednostka ε związana z γ. Ostatnia (dziewiąta) podjednostka oznaczona jest symbolem δ i znajduje się na zewnątrz F 1 .
Część błonowa syntazy ATP, zwana czynnikiem sprzęgającym F0, jest hydrofobowym kompleksem białkowym, który przenika przez błonę i ma wewnątrz dwa półkanały, przez które przechodzą protony wodoru. W sumie kompleks F0 zawiera jedną podjednostkę białka tego typu A, dwie kopie podjednostki B, a także 9 do 12 kopii małej podjednostki C. Podjednostka A(masa cząsteczkowa 20 kDa) jest całkowicie zanurzona w membranie, gdzie tworzy sześć przecinających ją odcinków α-helisy. Podjednostka B(masa cząsteczkowa 30 kDa) zawiera tylko jeden stosunkowo krótki obszar α-helikalny zanurzony w membranie, a jego pozostała część wyraźnie wystaje z membrany w stronę F 1 i jest przyczepiona do podjednostki δ znajdującej się na jej powierzchni. Każda z 9-12 kopii podjednostki C(masa cząsteczkowa 6-11 kDa) to stosunkowo małe białko zbudowane z dwóch hydrofobowych α-helis, połączonych ze sobą krótką pętlą hydrofilową skierowaną w stronę F 1 i razem tworzą pojedynczy zespół w kształcie walca zanurzonego w błonie . Podjednostka γ wystająca z kompleksu F 1 w stronę F 0 jest dokładnie zanurzona w tym cylindrze i dość mocno z nim związana.
Zatem w cząsteczce syntazy ATP można wyróżnić dwie grupy podjednostek białkowych, które można porównać do dwóch części silnika: wirnika i stojana. „Stator” jest nieruchomy względem membrany i zawiera sferyczny heksamer (αβ) 3 umieszczony na jego powierzchni oraz podjednostkę δ i podjednostki A I B kompleks błonowy F0. Ruchomy względem tej konstrukcji „wirnik” składa się z podjednostek γ i ε, które wyraźnie wystające z kompleksu (αβ) 3 połączone są z pierścieniem podjednostek zanurzonych w membranie C.
Zdolność do syntezy ATP jest właściwością pojedynczego kompleksu F 0 F 1, związaną z przeniesieniem protonów wodoru przez F 0 do F 1, w tym ostatnim, w którym znajdują się centra katalityczne przekształcające ADP i fosforan w cząsteczkę ATP . Siłą napędową działania syntazy ATP jest potencjał protonowy powstający na wewnętrznej błonie mitochondrialnej w wyniku działania łańcucha transportu elektronów.
Siła napędzająca „wirnik” syntazy ATP występuje, gdy różnica potencjałów pomiędzy zewnętrzną i wewnętrzną stroną membrany osiąga > 220 mV i jest zapewniana przez przepływ protonów przez specjalny kanał w F0, zlokalizowany na granicy podjednostek A I C. W tym przypadku szlak transferu protonu obejmuje następujące elementy strukturalne:
- Dwa niewspółosiowo położone „półkanały”, z których pierwszy zapewnia dopływ protonów z przestrzeni międzybłonowej do niezbędnych grup funkcyjnych F0, a drugi zapewnia ich wyjście do matrix mitochondrialnej;
- Pierścień podjednostek C, z których każdy w swojej środkowej części zawiera protonowaną grupę karboksylową, zdolną do przyłączania H + z przestrzeni międzybłonowej i uwalniania ich przez odpowiednie kanały protonowe. W wyniku okresowych przemieszczeń podjednostek Z, wywołany przepływem protonów przez kanał protonowy, podjednostka γ obraca się, zanurzona w pierścieniu podjednostek Z.
Zatem aktywność katalityczna syntazy ATP jest bezpośrednio powiązana z rotacją jej „wirnika”, w którym obrót podjednostki γ powoduje jednoczesną zmianę konformacji wszystkich trzech podjednostek katalitycznych β, co ostatecznie zapewnia funkcjonowanie enzymu . W tym przypadku w przypadku tworzenia ATP „wirnik” obraca się zgodnie z ruchem wskazówek zegara z prędkością czterech obrotów na sekundę, a sam taki obrót następuje w dyskretnych skokach co 120°, z których każdy towarzyszy tworzeniu się jednej cząsteczki ATP .
Bezpośrednia funkcja syntezy ATP jest zlokalizowana w podjednostkach β kompleksu koniugującego F1. W tym przypadku pierwszym aktem w łańcuchu zdarzeń prowadzącym do powstania ATP jest związanie ADP i fosforanu z centrum aktywnym wolnej podjednostki β, która znajduje się w stanie 1. Dzięki energii zewnętrznej źródła (prąd protonowy) w kompleksie F 1 zachodzą zmiany konformacyjne, w wyniku których ADP i fosforan zostają trwale związane z centrum katalitycznym (stan 2), gdzie możliwe staje się utworzenie pomiędzy nimi wiązania kowalencyjnego, co prowadzi do powstawanie ATP. Na tym etapie syntazy ATP enzym praktycznie nie potrzebuje energii, która będzie potrzebna w kolejnym etapie do uwolnienia ściśle związanej cząsteczki ATP z centrum enzymatycznego. Zatem kolejnym etapem działania enzymu jest to, że w wyniku zależnej od energii zmiany strukturalnej w kompleksie F 1, katalityczna podjednostka β zawierająca ściśle związaną cząsteczkę ATP przechodzi w stan 3, w którym połączenie ATP z centrum katalitycznym jest osłabiony. W wyniku tego cząsteczka ATP opuszcza enzym, a podjednostka β powraca do swojego pierwotnego stanu 1, który zapewnia cykl enzymu.
Praca syntazy ATP związana jest z mechanicznymi ruchami jej poszczególnych części, co pozwala zaliczyć ten proces do szczególnego rodzaju zjawiska zwanego „katalizą rotacyjną”. Tak jak prąd elektryczny w uzwojeniu silnika elektrycznego napędza wirnik względem stojana, tak ukierunkowany transfer protonów przez syntazę ATP powoduje rotację poszczególnych podjednostek czynnika koniugacji F 1 względem innych podjednostek kompleksu enzymatycznego, jak w wyniku czego to wyjątkowe urządzenie wytwarzające energię wykonuje pracę chemiczną - syntetyzuje cząsteczki ATP. Następnie ATP wchodzi do cytoplazmy komórki, gdzie jest zużywany na różnorodne procesy zależne od energii. Taki transfer odbywa się za pomocą specjalnego enzymu translokazy ATP/ADP wbudowanego w błonę mitochondrialną, który wymienia nowo syntetyzowane ATP na cytoplazmatyczny ADP, co gwarantuje bezpieczeństwo puli nukleotydów adenylowych wewnątrz mitochondriów.